第六节 免疫应答的调节
免疫应答是机体针对外来抗原产生的一种复杂的排斥过程,与其他生理系统相互配合,共同维持机体内环境的稳定。与其他生理过程一样,免疫应答也受到许多因素的影响和制约。首先,免疫应答受遗传基因的控制;受免疫系统内部各种因素的制约;还要接受宿主整体生理水平的调节。
一、TH细胞的调节作用
TH细胞不是一个均一的群体,活化后可分化成TH1、TH2和TH0三种类型。三个类型细胞免疫功能互有差别,尤其TH1、TH2和TH0三种类型。三个类型细胞的免疫功能互有差别,尤其TH1与TH2之间存在相互或促进的作用。因为TH是免疫应答的中心细胞,所以它的活性对整个免疫应答具有调节作用。
TH1产生IL-2、IFN和TNF,作用于各种免疫细胞。IL-2可诱导T细胞分裂增殖,增强细胞免疫应答;但IFN抑制抗体应答,抑制移植排斥反应和迟发型超敏反应。TH2分泌IL-4和IL-5等细胞因子,可诱导B细胞的增殖化,促进抗体产生(细胞因子的作用详见第五章)。由此可见,TH细胞在体液和细胞免疫应答中皆有调控作用,被视为免疫应答的中心调节作用途径。
二、抗体分子的调节作用
抗体是B细胞应答的效应产物,可反过来对特异性体液免疫应答产生反馈性抑制作用。抗体与相应抗原结合所形成的免疫复合物可结合到B细胞的表面Ig上,向胞内传入抑制信号,影响B细胞的活化和抗体的产生。例如将Rh抗体注射给刚分娩Rh+婴儿的Rh-母亲,则可阻止母亲产生Rh抗体,从而预防下一次妊娠时可能发生的新生儿溶血。
在对肿瘤的免疫应答中,某些抗体分子与肿瘤抗原结合后,不仅能介导任何免疫效应,还能阻止Tc对靶细胞的杀伤,实际上是抑制了细胞免疫效应,这类体称为封闭抗体(blockingantibody)。
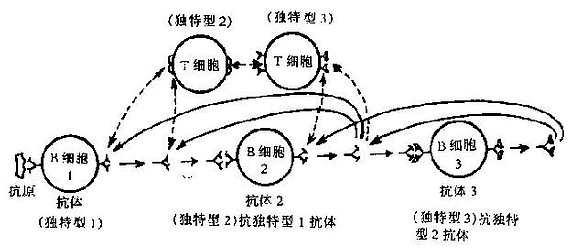
三、独特型网络的调节作用
与游离的Ig分子一样,B细胞的SIg分子高变区也存在着独特型标志。每一B细胞克隆的独特型标志能够被另一克隆B细胞的SIg所识别,构成一个相互识别的独特型网络;被识别的细胞受到抑制,而主动识别的细胞则活化。网络中的细胞可分成以下4组:①抗原反应细胞(antigen-rdactivecell,ARC),外来抗原与ARC结合,使细胞增殖并产生相应的抗体,构成网络的主体;②ARC抑制细胞,即抗独特型组(anti-idotypeset),可识别ARC的独特型标志,有抑制ARC反应的作用;③ARC激发细胞,为内影像组(internalimageset),该组细胞的独特位与外来抗原表位的结构相似,可模拟外来抗原对ARC构成刺激;④与ARC独特位相同的细胞,为非特异性平行组(unspecificparallelset),可被ARC激活细胞识别,刺激其对ARC激活细胞识别,刺激其对ARC的抑制作用。后三类细胞也分别抑制细胞和刺激细胞,以构成自己的网络(图7-6)。
图7-6独特型网络作用示意图
独特型网络调节的最终效应是抑制抗体的产生,使免疫应答终止。这一学说为免疫调节的研究开辟了新的领域,有关网络的精确机制及操纵方法尚需进一步研究。
四、免疫应答的整体调节
上述的免疫调节并非各自独立存在,而是相互影响,并且在整体上受神经-内分泌的调节,构成更加复杂的神经-内分泌-免疫调节网络。尽管免疫系统与神经系统和内分泌系统没有解剖学上的直接联系,但通过小分子介质可以沟通这三个系统。已有资料表明,免疫细胞可以表达某些神经递质的受体,而某些神经细胞上也发现有细胞因子的受体。虽然这些受体的确切作用尚未得到证实,但无疑这是系统间相互作用的物质基础。
现已证明雌激素、雄激素和皮质醇等可抑制免疫应答,而生长素、甲状腺素和胰岛素等则有免疫促进作用。乙酰胆碱、肾上腺素、去甲肾上腺素、多巴胺等神经递质对淋巴活性的影响都有报道,尤其内啡肽与脑肽对免疫应答的影响已受到重视,并证明它们可促进T细胞增殖和NK细胞的杀伤活性;而对抗体的产生则表现抑制作用。
大量的临床观察和实验研究表明,精神因素和条件反射对免疫应答也有显著的影响,但其作用途径及机制尚不清楚。
(尹学念)