第五节 免疫球蛋白的基因及抗体形成
免疫球蛋白反应的特异性和分子的多样性是受基因支配;一条肽链的C区和V区分别由C基因和V基因编码。任何一个B细胞都有3个独立的Ig基因簇:1个H链基因簇和2个L链基因簇(κ和λ),构成Ig的结构基因;在B细胞分化成熟过程中进行基因重排,进而转录与翻译,形成抗体。
(一)Ig基因的结构
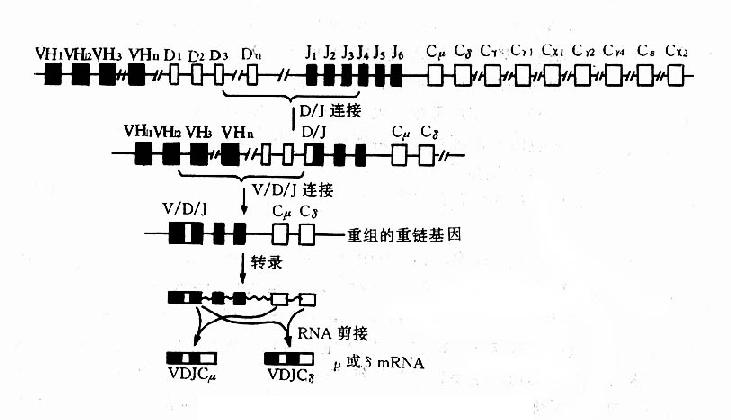
1.重链基因人类重链基因位于第14号染色体上,基因结构非常复杂,分为4个不连续的基因节段,从着丝点5'末端起依次为:可变区(VH)基因、多样性区(diversityregion,DH)基因、接合区(JH)基因和稳定区(CH)基因(图2-5)。
V区基因分成6个亚群,在2500kD的区域内排列有100~200个基因。某些大亚群如VHⅢ含有约25~30个基因,而某些小亚群如VHⅤ或VHⅥ仅含一个或几个基因。每个V基因由一个大的外显子和一个位于前导顺序后的内含子(约100~150bp)组成,前导顺序编码一种疏水肽,指引Ig肽链的转膜作用,V基因3末端是重组酶信号。在VH座内还有一些不具表达功能的假基因。
C基因结构约200kb,含有11个基因。第一个CH为Cμ,以后依次为:Cδ、Cγ3、Cγ1、φε1、Cα1、φγ、Cγ2、Cγ4、Cε、Cα2。其中φε1(φε2不在第14号染色体上)和φγ是两个假基因。除Cδ基因外,其他CH基因上游都有一个转换(S)顺序,负责H链的类转换。临床正常个体的CH座位内可有大片缺失,这种无免疫缺陷症状的个体可能是通过细胞选择在免疫应答中补偿这种基因缺失。
图2-5Ig重链基因及重组示意图
每个CH基因的外显子分别编码相应H链的功能区,由内含子将其隔开。如Cμ基因有4个外显子各自编码链C区上的Cμ1、Cμ2、Cμ3和Cμ4四个功能区。除上述主要的CH基因外,还有其他编码不同形式Ig分子的基因,例如分别编码分泌IgM和膜IgM的μs和μm基因;前者是Cμ45'端的外加部分,编码μs链C端20个氨基酸;后者位于Cμ基因下游,含2个外显子,共同编码μm链C端41个氨基酸。
V和C基因被中间的另两个基因节段分开,即D基因和J基因。J区有6个功能基因和3个假基因;D区的基因数目尚未确定,但至少不下20个。D和J基因参与重链V区的编码,负责其羧基端的一段氨基酸顺序。
2.轻链基因轻链的基因比重链的基因结构简单,仅有V区和J区而无D基因节段。κ链和λ链的基因互不相同(图2-6)。
人类κ链基因位于第2号染色体上。Cκ基因只有一个,邻近上游的J区座位内有5个Jκ基因,Vκ基因节段大约有80个Vκ基因,约一半以上可能是假基因。
λ链基因位于第22号染色体上。Cλ基因簇比Cκ基因复杂得多,至少有6个非等位基因,其中2个为假基因;每个功能Cλ基因前均有一个(或更多)相关的Jκ基因;对Vλ基因库目前所知甚少,其基因数目尚不清楚。
轻链的J基因参与V区肽链的编码,大约负责十几个氨基酸的顺序。
(二)Ig基因的重排
胚系状态的Ig基因,无论是重链基因还是轻链基因,都不能作为一个独立的单位进行表达,只有经过重排以后才能成为具有表达功能的基因。在成熟Ig基因的产生过程中,Ig基因的重排需遵循一定的顺序,先由V-J连接或V-D-J连接,然后由VJ或VDJ与C区基因连接(图2-5)。在重链,还可以发生类转换。
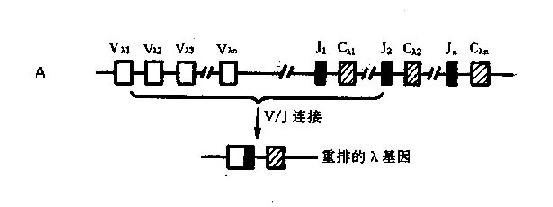

图2-6Ig轻链基因及其重组
A:λ基因;B:κ基因
1.V-J或V-D-J连接轻链的V-J连接和重链的V-J-D连接都是在DNA水平发生,均由重组酶介导。V-J或V-D-J的组合都是随机的;重组后的V-J编码轻链的V区,V-D-J编码重链的V区。
V-J基因重排是通过V区3'端和J区5'端旁的特殊顺序使V-J靠扰并提供酶切信息,实现V基因和J基因结合成为V-J基因单位。这种V-J重排是随机性的。V基因节段中任何一个V基因可与任何一个J基因重排结合。被结合的J基因上游的J基因丢失,下游的J基因保留。
V-D-J重排中,除V3'端和J5'端旁侧外,D两侧亦有上述特殊识别顺序在起作用。V-D-J连接中往往是DJ结合先于VD结合。与V-J连接一样,V-D-J重排也具有不精确性。还有严重排为功能性连接的VH被其中游胚系状态(未重排)的VH所替换。这可能是扩大基因容量和保证胚系状态的VH基因能全部利用的一种机制。
在B细胞成熟过程中,Ig基因存在重排的等级(hierarchy)现象。在多能造血干细胞分化发育成为幼稚B细胞(又称前B细胞)时,就发生V-D-J重排,开始表达H链,邻近J基因的Cμ自然随之表达,这是顺序优先的结果。由于Cμ基因和Cδ基因间的距离很短,两者可以同时得以转录;V-D-J在RNA水平既可与C结合,也可与Cδ结合,使IgM和IgD在单个B细胞上协同表达,而并非缺失性类转换。以后κ基因开始Vκ和Jκ重排,产生κ链。
2.重链类转换类转换是在DNA水平上V-D-J与CH基因连接由Cμ和Cδ转换成其他CH基因的过程,是其他CH基因上游的S顺序间发生重组的结果。S-S重组导致重组S顺序间的所有DNA基因丢失,例如Sμ与Sγ1间发生转换,则Cμ、Cδ和Cγ3基因及其侧面的顺序均一起丢失,使V-D-J连接由一个CH重新定位于另一个CH。类转换只变换Ig的类别,不改变抗体的特异性。
(三)Ig基因的表达及Ig分子的分泌
Ig的合成过程与一般蛋白质合成相似。在细胞内有表达功能的V-J或V-D-J基因单位重组完成后,与C基因簇一起被转录成初级RNA,经过加工剪接,去除内含子,生成mRNA,最后分别翻译成各种肽链,装配成Ig分子,分泌出体外。
Ig基因在表达时存在等位排斥(allelicexclusion)和同型排斥(isotypicexclusion)现象,可能是V-D-J连接或V-J连接的不精确性所造成的结果,以致许多重排无转录产物。一个B细胞不会同时表达κ链和λ链,称同型排斥。κ基因重排总发生在λ基因重排之前,当Vκ-Jκ重排形成有表达功能的基因后,λ基因重排即被抑制;在λ链产生细胞内,常有κ基因缺失。象其他的基因一样,Ig基因的表达过程中也有启动子与增强子来启动和调节基因的转录。
B细胞在接受抗原刺激后迅速分化增殖,除一部分分化记忆细胞外,其余分化为浆细胞。浆细胞在内质网和多聚糖体均显著增加,大量合成Ig分子。合成L与H链的粗面内质网多聚核糖体是不同的。L链在190~200S的多聚核糖体(含4~5个核糖体)上合成,H链在270~300S的多聚核糖体(含11~18个核糖体)上合成。作为一条完整的多肽链,它们从一个起始点(N端)开始(向C端)依次合成。游离的L和H链少数在多聚核糖体上就有非共价结合或共价结合,大部分转移至内质网的贮池中,并装配成完整的Ig分子,然后依赖N端疏水性前导顺序进入高尔基复合体,再分泌至细胞外。在此移动过程中糖残基通过结合在膜上的糖转化酶按一定顺序逐步加到Ig分子上。
(四)抗体分子的多样性
一个机体何以能产生多达106~108种具有不同抗体特异性的Ig分子,其机制至今虽未完全清楚,但从基因的结构组成及重排中可找到一些答案。众多V区基因和一个或少数几个C区基因不连续地排列在染色体上,它们在DNA水平随机地结合是Ig分子多样性的基础,而体细胞突变又可增大V区的库容。
多样性程度可以通过Ig基因在染色体内重组时V-J与V-D-J的乘积来计算:当100个Vκ和5个Jκ重组时所产生的多样性至少是100×5=5×102个;V-D-J重排时100个VH与10个DH和6个JH连接所的生的多样性至少有100×10×6=6×103。同时连接这些基因时还会发生不精确性而使多样性增加,因而由κ链和H链组成的抗体分子的多样性最少有5×102×6×103=3×106之多。另外,在V-J、V-D-J连接过程中发生的碱基缺失和插入又扩大了多样性的程度。
(尹学念)