3.3 脂类的生化代谢
3.3.1脂类的消化和吸收
唾液中无消化脂肪的酶,胃液中虽含有少量的脂肪酶,但成人胃液酸度很强,不适于脂肪酶的作用,故脂肪在成人口腔和胃中不能消化。婴儿胃液的PH在5左右,奶中脂肪已经乳化,故脂肪在婴儿胃中可消化一部分。脂肪的消化主要在小肠内进行。食糜通过胃肠粘膜产生的胃肠激素刺激胰液和胆汁的分泌,并进入小肠。胆汁中的胆盐是强有力的乳化剂,脂肪受到胆盐的乳化,分散为细小的脂肪微粒,有利于和胰液中的脂肪酶充分接触。
胰液中的胰脂肪酶能将部分脂肪完全水解为甘油和游离脂肪酸,但有一半的脂肪仅能局部水解为甘油二酯或甘油一酯。因为胰脂肪酶能特异地和连续地作用于甘油三酯的1和3位置,开始解脱一个脂肪酸,形成甘油二酯。然后,再解脱一个脂肪酸,形成甘油一酯。胰脂肪酶对甘油三酯的水解率和其脂肪酸链的长短有关,不饱和脂肪酸比饱和脂肪酸易于水解。还有小部分的脂肪完全不水解。
脂肪的水解产物游离脂肪酸和甘油一酯、甘油二酯进入肠粘膜细胞内,在滑面内质网上重新合成与体内脂肪组成成分相近的甘油三酯。新合成的甘油三酯的组成和构型适宜于以后的代谢。新合成的甘油三酯在粗内质网上与磷脂、胆固醇、蛋白质形成乳糜微粒,经肠绒毛的中央乳糜管汇合入淋巴管,通过淋巴系统进入血液循环。水解产物甘油因水溶性大,不需胆盐即可通过小肠粘膜经门静脉而吸收入血液。完全未被水解的脂肪亦能以乳胶微粒的形式直接进入肠粘膜细胞,在内质网上合成的乳糜微粒再由淋巴系统进入血液循环。因此,动物和植物脂肪几乎完全吸收。食后2h,可吸收24~41%,4h后吸收53~71%,6h后吸收68~86%,12h后吸收97~99%。
影响脂肪吸收的因素:
(1)脂肪的熔点脂肪的熔点会影响其吸收。例如,羊脂的熔点为44~55℃,其吸收率为85%;而椰子油的熔点为28~33℃,其吸收率为98%。一般说来,植物油的熔点较低,所以较易吸收。这是因为进入十二指肠中的脂肪应该是液态,这样才能乳化。脂肪的熔点比体温越高,就越难于乳化,所以也就越不容易消化吸收。
(2)脂肪摄取量因为脂肪吸收比较慢,小量食入时吸收率高。大量时有不少排泄掉,吸收率低。
(3)年龄1岁内的婴儿脂肪吸收率低,常易发生消化不良。老年人脂肪的吸收和代谢都比年轻人慢。
(4)脂肪酸组成一般来说,含短链脂肪酸的脂肪其吸收比长链的为快。含奇数碳链脂肪酸的脂肪,其吸收比偶数的为慢。棕榈酸在甘油第2位的脂肪(如猪油、人乳),其吸收比棕榈酸在其他位置或在第2位的其他脂肪酸的脂肪(牛油、羊油、牛乳)来得好。
(5)钙脂肪吸收时,虽然需要一定量的钙,但如钙量过高时,则脂肪吸收反而下降。特别是含月桂酸、豆蔻酸、软脂酸和硬脂酸等熔点高的脂肪。而含油酸和亚酸这些不饱和脂肪酸浓度高的脂肪,钙不影响其他吸收。
钙干扰饱和脂肪吸收的机理是由于形成难溶解的饱和脂肪酸钙。
3.3.2磷脂的消化和吸收
卵磷脂在小肠内由四种酶进行分解,由胰腺分泌的磷脂酶A原,受胰蛋白酶激活成磷脂酶A,在胆盐和Ca2+存在下,作用于卵磷脂,释出一个脂肪酸,产生溶血卵磷脂。它有溶血作用。
磷脂酶B作用于卵磷脂,释出二分子脂肪酸,产生α-甘油磷酸胆碱。溶血卵磷脂亦可受磷脂酶B的作用,释出一分子脂肪酸后,生成α-甘油磷酸胆碱。
最后,甘油磷酸酶和胆碱磷酸酶分别作用于α-甘油磷酸胆碱,完全水解成甘油、磷酸及胆碱。
脑磷脂和磷脂酰丝氨酸的分解过程与卵磷脂相似。
除脂肪酸外,磷脂的消化产物大多数是水溶性的,在肠道内易于吸收。
小部分磷脂在胆盐的协助下,混合在乳胶微粒内,在肠内可以不经消化而能直接吸收。但大部分磷脂仍需水解后才被吸收。吸收的磷脂水解产物,也可以在肠壁重新合成完整的磷脂分子再进入血液中。
3.3.3胆固醇的消化和吸收
食物中所含的胆固醇,一部分是与脂肪酸结合的胆固醇酯,另一部分是游离状态的。胰液和肠液中均含有胆固醇酯酶,在肠道内催化胆固醇脂水解,产生游离的胆固醇和脂肪酸。
胆固醇为脂溶性物质,故必须借助胆盐的乳化才能在肠内吸收。但是吸收的胆固醇约有三分之二在肠粘膜细胞内经酶的催化重新酯化,形成适合体内需要的胆固醇酯。再与部分未酯化的游离胆固醇、磷脂、甘油三酯及由肠粘膜细胞合成的脱辅基蛋白一起形成乳糜微粒,经淋巴系统进入血液循环。因此,淋巴和血液中的胆固醇大部分以胆醇酯的形式存在。
影响胆固醇吸收的因素:
(1)食物胆固醇在摄食1、3或6g胆固醇后,其吸收率分别为60、40、30%,即吸收率随着摄食量的增加而递减。这是因为增加胆醇吸收的同时,发生两个代偿机制,即胆固醇排泄增加和体内合成减少。
(2)食物脂肪和脂肪酸食物中的脂肪和脂肪酸具有提高胆固醇吸收的作用。这是由于:①乳糜微粒中的胆固醇主要是胆固醇酯,脂肪和脂肪酸可以在肠粘膜中供给胆固醇以再酯化所需要的脂酰基,从而有利于胆固醇吸收;②高脂肪膳食具有促进胆汁分泌的作用。而胆汁中的胆汁盐能促使胆固醇形成微粒,有助于胆固醇的吸收。
(3)植物固醇各种植物固醇,如豆固醇、谷固醇等,不仅其本身吸收很差,而且还能抑制胆固醇的吸收。有人认为这可能是因为:①植物固醇的分子结构与胆固醇极为相似,竞争性抑制肠内胆固醇酯的水解,以及肠壁内游离胆固醇的再酯化,促使其由粪便中排泄;②植物固醇竞争性地占据微粒内胆固醇的位置,影响胆固醇与肠粘膜细胞接触的机会,从而妨碍其吸收。
(4)其它食物中不能被利用的多糖,如纤维素、果胶、琼脂等容易和胆汁盐结合形成复合物,妨碍微粒的形成,故能降低胆固醇的吸收。此外,肠细菌能使胆固醇还原为不易吸收的粪固醇。因此,长期服用广谱抗生素的病人,常能增加胆固醇的吸收。
3.3.4脂肪的合成
脂肪合成有两条途径:一是利用食物中的脂肪转化而成人体脂肪;另一是将糖转变为脂肪,这是体内脂肪的主要来源。脂肪组织和肝脏是体内脂肪合成的主要场所。合成脂肪的原料是磷酸甘油和脂肪酸。
磷酸甘油是由糖代谢的中间产物磷酸丙糖还原而成,或从食物中消化吸收的甘油在甘油激酶的催化下,与ATP作用而生成磷酸甘油。
脂肪酸用于脂肪合成之前需经活化,即在脂肪酰CoA合成酶的催化下,与辅酶A、ATP作用生成脂酰辅酶A。二分子脂酰辅酶A经过脂酰转移酶的催化。将脂酰基转移到α-磷酸甘油分子上,生成α-磷酸甘油二酯,又称磷脂酸。后者经磷脂酸酶作用,脱去磷酸后再与另一分子脂酰辅A在甘油二酯转脂酰酶的作用下,结果就生成脂肪。
3.3.5磷脂的合成
体内磷脂一部分是直接由食物中来,另一部分是在各组织细胞内,经过一系列酶的催化而合成。磷脂的种类很多,现仅将甘油磷脂和神经磷脂的合成简述如下:
(1)甘油磷脂的合成甘油磷脂主要是在微粒体内合成,其原料为磷酸、甘油、脂肪酸、胆碱或乙醇胺等。其中必需脂肪酸由食物供应,其他原料可在体内合成。蛋白质分解产生的甘、丝及蛋氨酸即可作原料。甘氨酸在体内经亚甲基四氢叶酸作用变为丝氨酸,再脱羧变乙醇胺,由S-腺苷蛋氨酸供给甲基而变为胆碱。胆碱和ATP在胆碱磷酸激酶催化下生成磷酸胆碱,再和胞苷三磷酸(CTP)在胞苷酸转移酶作用下变为胞苷二磷酸胆碱(CDP-胆碱),他和甘油二酯在磷酸胆碱转移酶催化下脱掉胞苷一磷酸(CMP),就形成α-卵磷脂。
脑磷脂的合成与卵磷脂的合成过程基本相似,不同的的只是以乙醇胺代替胆碱。
磷脂还可以从另一条途径合成,即α-磷酸甘油二酯先与CTP作用生成CDP-甘油二酯,再与丝氨酸反应生成磷脂酰丝氨酸,后者直接脱羧即生成脑磷脂。脑磷脂甲基化即可生成卵磷脂。
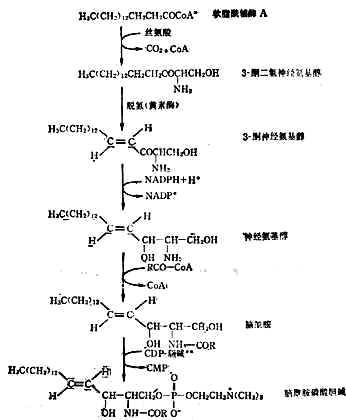
(2)神经磷脂的合成人体内的神经磷脂种类很多,现仅将脑酰胺为中间产物的一种合成过程列于图3-4。
3.3.6胆固醇的代谢
(1)胆固醇的合成人体内的胆固醇一部分(约40%)是由动物性食物中来,称为外源胆固醇;一部分是由体内组织细胞自行合成,称为内源胆固醇。
人体内几乎所有组织都具有合成胆固醇的能力,但合成的速率和总合成量并不一致,肝脏不仅合成快,而且合成量也最多,是人体合成胆固醇最活跃的场所,其次是小肠。
合成的原料可来自乙酰辅酶A。前后经30多步酶促反应,全部过程在细胞质内进行。
图3-4 脑酰胺磷酸胆碱的合成途径
为了储存和转运的目的,游离胆固醇和长链脂肪酸结合成胆固醇酯。组织中的胆固醇是在胆固醇酰基转移酶的作用下,接受脂酰CoA的脂酰基形成胆固醇脂。血浆胆固醇的酯化是在血浆中的卵磷脂胆固醇基转移酶(LCAT)催化下进行的。
(2)胆固醇合成的调节在调节胆固醇的合成中,β-羧-β-甲戊二酸(HMG)-CoA还原酶具有决定性意义,任何因素改变还原酶的活性时,则显著影响体内胆固醇的合成。食入高胆固醇后,当胆固醇含量升高时,可反馈抑制肝脏胆固醇的合成。此作用主要是还原酶活性下降之故。这种负反馈机制只发生于大鼠和其它动物的肝脏,而在胃肠道却没有发现。至于人体,外源性胆固醇不能降低肝外组织的胆固醇合成。其对肝脏的影响,仍有不同的意见。因此,大量摄食胆固醇后,血浆胆固醇仍有一定程度升高。
胆固醇合成速率在昼夜之间可相差4~5倍,午夜时合成最快,上午10时左右最慢,这与还原酶活性的变化是一致的。
还有许多激素也对此酶有影响,如肾上腺素和去甲肾上腺素能促进此酶的合成,因此使胆固醇合成增加。甲状腺素能增加组织对儿茶酚胺的敏感性,而后者又增加HMG-CoA还原酶的合成。但甲腺素又能增加胆固醇的分解和排出,而后者的作用却大于前者,结果使血清胆固醇降低。故甲亢病人常见血浆胆固醇降低,而甲状腺机能减退病人则血浆胆固醇增高。cAMP也有抑制胆固醇合成的作用,于是有人推测有些激素的作用是通过cAMP的变化而实现的。
肝脏内胆固醇的合成还受脂肪代谢的影响。当脂肪酸动员加强,不仅血甘油三酯升高,胆固醇合成也明显增强。运动能使血浆游离脂肪酸含量减少,从而使胆固醇合成缓慢。多不饱和脂肪酸可在体内转变成前列腺素,有人认为前列腺素可通过cAMP-蛋白质激酶系统而使HMG-CoA还原酶失活,或通过抑制脂肪动员,降低血浆游离脂肪酸而使肝中胆固醇合成减少。这可能是多不饱和脂肪酸降低血浆胆固醇的一个重要原因。
糖、脂肪和蛋白质分解而产生的乙酰CoA主要经三羧酸循环彻底氧化,但过量的乙酰CoA不仅可以合成脂肪,也可在微粒体酶系的作用下大量生成HMG-CoA。因此,HMG-CoA还原酶的酶促反应随之加快,胆固醇合成增加。
(3)胆固醇的转化胆固醇在体内能转变成一系列有生理活性的重要类固醇化合物:①转变为胆汁酸。胆固醇在肝脏内受7α-羟化酶的催化生成7α-羟胆固醇,后者经一系列反应转变为胆汁酸。这是胆固醇的主要去路,人体内约有80%的胆固醇可以在肝脏中转变为胆汁酸,其中主要是胆酸、脱氧胆酸和鹅脱氧胆酸。胆酸再与甘氨酸或牛磺酸结合成胆汁酸。胆汁酸以钠盐或钾盐的形式存在,称之为胆盐。它们对脂类的消化吸收起重要作用;②转变为维生素D3。在肝脏和肠粘膜细胞内,胆固醇可转变为7-脱氢胆固醇。后者经血液循环运至皮肤,再经紫外光照射,7-脱氢胆固醇可转变为维生素D3。维生素D3能促进钙磷吸收,有利于骨胳生成;③转变成类固醇激素。胆固醇在肾上腺皮质细胞内转变成肾上腺皮质激素,如醛固酮,皮质醇。再卵巢内可转变为孕酮与雌激素。在睾丸内可转变为睾酮。
(4)胆固醇的排泄部分胆固醇可以在组织内(主要是肝脏)还原,生成二氢胆固醇,与胆固醇一起分布于全身各组织。体内胆固醇由肝脏排入胆汁,随胆汁进入肠腔。一部分通过肠肝循环重新吸收入肝脏。一部分在肠道被经细菌作用后转变为粪固醇。未被吸收的二氢胆固醇以及很难吸收的的粪固醇统称为中性粪固醇,随粪便排出体外。
此外,尚有少量胆固醇和二氢胆固醇以皮脂形式由皮肤排出;小部分胆固醇和胆固醇脂随表皮细胞脱落,一起排出体外。
3.3.7脂蛋白的代谢
血浆脂蛋白包括乳糜微粒、极低密度脂蛋白、低密度脂蛋白和高密度脂蛋白。它们的代谢简述如下:
(1)乳糜微粒(CM)的代谢CM的90%是甘油三酯,其余为磷脂、蛋白质和胆固醇。食物脂肪的水解产物经小肠吸收,并在小肠粘膜上皮细胞滑面内质网中重新合成甘油三酯。这些甘油三酯和从食物吸收的磷脂、胆固醇结合,同时与糙面内质网合成的脱辅基蛋白(apo)B和A-I形成原始的CM。它经过高尔基体加工后进入淋巴系统,称为淋巴CM。淋巴CM到达血液后,在组成成分和结构方面都发生一些改变。它主要包括接受由HDL转移来的apoC,同时还向血浆中释放磷脂和吸收胆固醇,这种CM遂变为成熟CM。
当血液经过脂肪组织、肝脏、肌肉等的毛细血管时,经管壁脂蛋白脂酶的作用,可使CM中的甘油三酯水解成脂肪酸和甘油。这些水解产物的大部分则进入细胞被利用或重新合成脂肪而储存。由于CM失去中心部分的甘油三酯而逐渐变小,这种CM称为CM残余。此时CM表面上的脱辅基蛋白、磷脂、胆固醇都脱离而移到HDL上。此种作用进行得很快,所以正常人空腹血浆几乎不易检出CM。有人认为CM残余可能在肝中转变为LDL。
(2)极低密度脂蛋白(VLDL)的代谢VLDL主要由肝实质细胞合成,其合成及分泌过程与小肠粘膜上皮细胞合成CM的过程基本相似。VLDL主要成分也是甘油三酯,但磷脂和胆固醇的含量比CM的多。其蛋白质部分除apoB以外,还有apoCⅠ、Ⅱ、Ⅲ,apoE,少量的apoAⅠ、Ⅱ和apoD。
肝细胞合成VLDL的甘油三酯,其来源是由糖在肝细胞中转变而来,也可由脂库中脂肪动员出来的游离脂肪酸在肝细胞的滑面内质网中重新合成。所以,VLDL是转运内源性脂肪的主要运输形式。此外,糙面内质网合成apoB,再与质膜的磷脂形成复合体,最后在高尔基体内结合成VLDL。它所含的胆固醇酯的来源不十分明了,可能是HDL输送来的。
VLDL和CM一样,经肝外脂蛋白脂酶的作用,使其中的甘油三酯水解成脂肪酸和甘油,被细胞利用或重新合成甘油三酯而储存。由于甘油三酯减少,胆固醇相对地增多,VLDL的结构遭受破坏。ApoC脱离这种已经改变的脂蛋白,使脂蛋白脂酶的作用停止。与此同时,LCAT酶被apoA-Ⅰ和apoC-Ⅰ激活,开始发挥催化作用,将一部分卵磷脂的β-脂酰基转移至胆固醇分子上,前者成为溶血卵磷脂,后者成为胆固醇酯。溶血卵磷脂的极性较大,便释放于血浆中。胆固醇没有极性,于是转移到脂蛋白的核心部分。经过上述变化,脂蛋白的体积比原来缩小,其所含的总胆固醇相对地增多,甘油三酯和apoC相对减少,但仍能维持一定的球形,通常把这种VLDL残骸称为中间低密度脂蛋白(ILDL)。
ILDL的成分是:apo15%,胆固醇29%~33%(其中75%为酯型),磷脂17%,甘油三酯35~39%。与VLDL比较,它的酯型胆固醇含量较高,而甘油三酯含量较低。
(3)低密度脂蛋白(LDL)的代谢VDLD水解产物ILDL到达肝脏,与肝细胞膜上的ILDL受体结合,ILDL遭受分解,释放出“多精肽”与其它脂类,最后变成LDL。人体的VLDL不是全部的话,亦是绝大部分转变为LDL。
与VLDL相比,LDL的胆固醇酯增多,apoB几乎没有减少,甘油三酯显著下降,而除apoB以外的蛋白质、磷脂等成分都除去了。所增加的胆固醇酯,可能是由于HDL的胆固醇经LCAT的作用而酯化,并转移到LDL上来的。
LDL的分解主要在肝外实质细胞中进行。细胞表面上有LDL的特异的受体,LDL通过它的apoB的正电荷精氨酸残基与此受体结合。结合的LDL在细胞表面的凹陷结构内为表面膜包成小泡,被摄入细胞内。含LDL的小泡与细胞内的溶酶体融合,LDL的蛋白质被溶酶体内的蛋白酶水解为氨基酸。酸性脂肪酶将胆固醇酯分解为游离胆固醇和脂肪酸。这种游离胆固醇能抑制能微粒体内HMG-CoA还原酶的活性,因而阻止细胞内胆固醇的合成。同时,又使微粒体内的LCAT酶活化,使胆固醇转变为适于储存的胆固醇酯(图3-5)。当细胞内游离胆固醇水平增高以及胆固醇酯开始堆积时,LDL受体的合成受到抑制,LDL吸收率下降。这样能维持血浆中和细胞内胆固醇浓度的平衡。

图3-5 人体成纤维细胞内LDL降解步骤
在正常人体内每天降解的LDL为总量的45%,其中约有三分之二是通过LDL受体途径进行的。此外,还有另一个降解途径,就是通过清除细胞(Scavenger cells)的吸入和降解LDL。这种细胞是从事于非特异性的胞饮作用。由这条途径所降解的血浆LDL是个常数(15%)。
(4)高密度脂蛋白(HDL)的代谢在肝脏或小肠内,CM经脂蛋白脂酶的作用,将甘油三酯分解,水解产物及其表层的磷脂、游离胆固醇和apoA离开CM而形成双层脂类组成的颗粒,这就是新生HDL,它呈碟形。这种新生HDL进入血液后,其表面被来自周围组织和其它脂蛋白的大量游离胆固醇所占据。此时,这些颗粒成为LCTA的最适底物,在LCTA催化下,使表面的游离胆固醇转变为胆固醇酯,并由颗粒表面转入颗粒核内。因核内脂类含量剧增,内压升高,使磷脂双层压变为单层。原有的碟形排列消失,形成一种富含胆固醇酯的成熟球形颗粒。这样,富含游离胆固醇的碟形新生HDL就转变为富含胆固醇酯的球形成熟HDL。
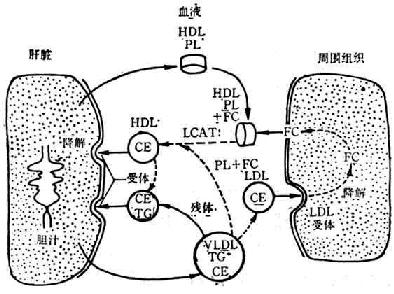
FC-游离胆固醇 CE-胆固醇酯 PL-磷脂 TG-甘油三酯
图3-6 HDL代谢特点
由于HDL代谢的特点,它能从周围组织转运胆固醇到肝脏进行降解排泄(图3-6)。这样,能防止胆固醇沉积在血管壁上,甚至已经沉积的胆固醇,亦能由HDL予以转移,可以防止并有可能消除动脉粥样硬化的形成。
肝脏和小肠是HDL的主要降解部位。血液中的成熟HDL,和细胞膜上的受体结合,进入细胞内,由溶酶体予以降解。分解出来的胆固醇一部分不变,一部分转变为胆酸而从胆汁排出。少量的HDL亦可在肾脏、肾上腺、卵巢等器官内降解。