第二节 常见遗传性疾病的发生与基因诊断
一、血红蛋白分子病
血红蛋白由四种珠蛋白肽链组成,它们分别是α、β、δ和γ肽链,由它们不同的组合组成各种血红蛋白。除α肽链的基因位于16号染色体外,其余β、δ、γ肽链的基因连锁在11号染色体。血红蛋白异常导致的分子病就是珠蛋白结构基因的DNA分子结构异常所致。
⒈点突变 DNA分子的碱基转换或颠换造成单个碱基替代,导致三联体遗传密码改变,使得相应表达翻译的肽链中氨基酸发生改变而导致蛋白质结构改变,最终引起该蛋白的生理功能发生改变。如β链第6位应是谷氨酸,其对应的碱基密码是GAA,当颠换成GUA时,表达的氨基酸改为缬氨酸,当转换成AAA时,表达的氨基酸改为赖氨酸。在目前发现的血红蛋白异常中,绝大多数属于这种类型。
⒉终止密码子突变 mRNA翻译蛋白质的终止密码有UAA、UAG、UGA。当终止密码子发生点突变,如UAA转换为CAA时,终止密码翻译为谷胺酰胺,肽链无法终止导致肽链延长。相反,肽链中途某一密码突变终止密码时,肽链就提前终止变短。如在中国人中发现有β珠蛋白基因突变,使转录mRNA密码子17由AAG→UAG,使翻译的β珠蛋白过早终止,造成β链过短,而无正常功能,导致珠蛋白生成障碍性贫血发生。
⒊移码突变 由于DNA分子三联体密码子之间是无标点符号的,因此当DNA的碱基序列中缺失或插入一个核苷酸,就相当于多了或少了一个碱基,使得在突变位置以后的三联体密码子均依次发生变化,包括终止密码。所以移码突变可以是肽链的氨基酸发生改变,也可是肽链长短发生改变。而且这种突变位置越靠近翻译起始端(5′端)其后果越严重。如在中国人中发现有β珠蛋白基因转录的密码子41-42产生缺失,造成β链合成提前终止,这种β链不稳定而导致β珠蛋白生成障碍性贫血发生。
突变类型检测可用核酸测序,人工合成寡核苷酸探针作产前诊断。前者多用于研究,后者可应用于临床。现可利用PCR法扩增突变区域,再行测序或杂交分析,大大提高了灵敏度和特异性,可省去目的基因的克隆,方法也得到一定的简化。
⒋密码子缺失和插入它与移码突变不同的是生殖细胞在减数分裂时,染色单体发生错配或不等交换,造成在基因DNA分子中,缺失或插入的核苷酸不是一个,而是一部分,即多了或少了一部分密码子。当然其肽链也相应缺失或插入一个以上的氨基酸。如珠蛋白生成障碍性贫血发现二种类型,一种为右侧缺失型,缺失了涉及α2和α1基因3.7kb的片段,而左侧缺失型则缺失了α2及左侧区域4.2kb的片段。(α2基因排列在左,α1基因排列在右)。该类变异可用Southern blot法,RFLP法加以诊断,参见诊断分子生物学基本技术有关章节。
⒌融合基因由于减数分裂时不同珠蛋白肽链的基因之间发生不等交换,结果造成某一珠蛋白基因同时融合两种不同珠蛋白基因的部分碱基,由此合成的肽链也含有两种不同珠蛋白的氨基酸序列。
血红蛋白病的诊断可根据临床症状,如溶血性贫血等现象。有些没有临床症状,须依赖实验室诊断。在细胞水平可以通过血液学检查,蛋白分子水平也可通过电泳,热稳定试验等方法检查。
血红蛋白病的治疗目前尚无根治办法。该病的起因是基因突变所致,其治疗有待于基因工程技术来矫正基因的缺陷,即用基因治疗的方法加以根治。但要进行基因治疗,必须建立完善基因诊断的技术,这是医学检验工作者面临的新课题。目前基因诊断技术可利用核酸探针进行分子杂交或进行核酸测定来分析基因突变和突变的位置。由于mRNA分子直接转录了DNA分子上基因的信息,又直接指导蛋白质合成,在细胞内又有一定的量,故易于分离纯化和分析,因此对内源性基因的分析,其分析对象多为mRNA分子。mRNA分析最常使用的分子杂交技术有斑点杂交和Northern blot技术以及逆转录PCR(RT-PCR)技术。
二、先天性代谢缺陷病
(一)苯丙酮尿症
苯丙酮尿症(PKU)因患者尿中含大量苯丙酮酸而得名病因是患者肝缺乏苯丙氨酸羟化酶,使由食物摄入体内的苯丙酮酸不能正常代谢为酪氨酸,导致血清中苯丙氨酸浓度升高,可高达50-100mg/dl(正常参考值为1-3mg/dl)。大量的苯丙氨酸使旁路代谢活跃,经苯丙氨酸转氨酶作用生成苯丙酮酸。
苯丙氨酸羟化酶是在肝细胞中合成的,即在肝细胞中专一表达而在胎儿的绒毛细胞或羊水细胞中并不表达,给PKU的产前诊断带来了困难。
由于诊断分子生物学技术的发展,1983年胡流清等人完成了人苯丙氨酸羟化酶cDAN探针的制备。他们利用探针和Southern blot技术进行分子杂交,通过限制性片段长度多态性(restriction fragment-length polymerphism ,RFLP)分析实现了PKU的产前基因诊断。他们先用MSP-I限制性内切酶消化正常人基因组DNA,经核酸电泳后,将DNA片段转移到一张硝酸纤维膜(或尼龙膜)上,再用32p标记的人苯丙氨酸羟化酶cDNA探针杂交后,再经放射自显影技术显示正常人群有23kb和19kb两种限制性片段长度。同时他们调查了一个PKU家系,进行比较发现,同样用Msp-I内切酶消化,父母同时有23kb和19kb DNA,患儿只有19kb片段。说明在该家系中,苯丙氨酸羟化酶突变基因是与19kb片段连锁的。连锁简而言之就是指同一条染色体上的基因联合遗传的现象。因此在该家系中的第2胎,如果只出现19kb DNA片段将是患儿,而同时出现23kb和19kb DNA片段或只出现23kb DNA片段的都是正常个体,而且只出现23kb DNA片段者不携带突变基因,不是突变基因的携带者。正常人出现19kb仅代表蛋白质和酶的多态。PKU家系的患儿为19kb,正常儿为23kb说明该PKU家系的基因突变与19kb连锁。(表15-2)
表15-2 人苯丙氨酸羧化酶基因核酸电泳与cDNA探针杂交自显影
| 正常人多态 | PKU家系 | |||||
| 父 | 母 | 患儿 | 正常胚胎 | |||
| 23kb | - | - | - | - | - | |
| 19kb | - | - | - | - | ||
1986年胡流清等进一步证明苯丙氨酸羧化酶的表达异常是基因点突变所致,并确定了突变点位置,并由此设计了正常和突变的各含21个核苷酸的寡核苷酸探针。
↓
正常探针5′TCCATTAACAGTAAGTAATTT3′
突变探针5′TCCATTAACAATAAGTAATTTT3′
↑
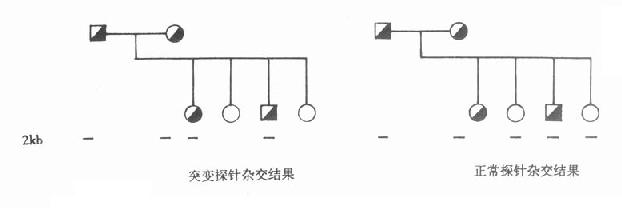
用这两个探针分别与PKU家系成员的DNA杂交,证实与正常探针杂交者为正常个体,与突变探针杂交者为患者,同时可与两个探针杂交者为正常个体,但携带突变基因。此法已成功用于产前诊断。(图15-1)
图15-1 正常与突变苯丙氨酸羟化酶探针与PKU家系成员的DNA杂交结果
杂交技术由于费时、操作复杂而难以在基层实验室推广。现在多用聚酶链反应(PCR)扩增技术进行基因诊断,或在PCR基础上再进行探针杂交,在提高灵敏度的同时又提高特异性,而且快速、经济、简便实用。
(二)嘌呤代谢紊乱与痛风
尿酸是嘌呤核苷酸分解代谢的重要产物,血和尿中尿酸浓度的检测是嘌呤代谢紊乱的重要化学指标。可由经典的磷钨酸还原法和高度特异的尿酸酶等化学方法检测。
健康成年男子每天约生成尿酸600-700mg,其中60%-70%经肾排出,剩余约200mg排入肠道由细菌降解。体内尿酸池维持在1200mg尿酸水平。肾排出尿酸的机制是由于尿酸分子量小(168u),可全部经肾小球滤过,但至少有98%被近曲小管重吸收,再经远曲小管主动分泌,因此随尿排出的尿酸主要由肾小管所分泌。肾每天约排出尿酸400-500mg(2.4-3.0mmol/L),相当于肾小球原滤液中含尿酸的4%-5%。
在血浆pH为7.4时,尿酸几乎完全以单钠尿酸盐的形式存在,溶解度有限,约为0.42mmol/L(7.0mg/dl);而尿酸的溶解度更低,在pH5的尿液中,比尿酸难溶20倍。健康成年男子的血清尿酸浓度约0.3mmol/L,女子约低20%。血清尿酸水平随年龄增加而增高,男性比女性更明显,可达0.46mmol/L。血清尿酸超过0.42mmol/L为高尿酸血症(hyperuricemia)。在血清尿酸浓度超过尿酸盐的溶解度时,就有可能尿酸钠的针状结晶沉淀于关节、肌腱、韧带、肾锥体的间质组织等软组织。足够数量的尿酸盐结晶可引起急性炎症反应,如沉积于关节腔,形成急性关节炎。这是由于尿酸盐结晶被白细胞吞噬后,破坏溶酶体膜,使膜内酶释放损伤白细胞和周围组织,引起关节炎症状。表现为关节剧烈疼痛,反复发作,此即痛风(gout)。沉积于软组织的结晶称为痛风石,周围发生炎症反应则构成痛风结节。原发痛风可由于遗传所致的核苷酸代谢中某一种酶表达异常所致,继发性痛风则继发于多种疾病。继发性病因可能有大量服用葡萄糖、果糖与甘露糖,使体内嘌呤合成增加;或多发性骨髓瘤、红细胞增多症、恶性贫血、牛皮癣和广泛转移的恶性肿瘤,使核蛋白转换率增快,而尿酸生成过多;或细胞毒药物或放射治疗时,核酸分解亢进使尿酸盐进一步增高。
先天性遗传原发通风病因是次黄嘌呤-鸟嘌呤磷酸核糖转移酶(HGPRT)缺失所致,部分缺失时,临床表现为尿酸过多的痛风特征;当完全缺失时,表现为高尿酸血症、精神发育迟缓等特征的自毁容貌综合征。
HGPRT蛋白质分子为相同亚基的四聚体,每个亚基由217个氨基酸残基组成。HG-PRT基因定位于X染色体长臂远端,因此该病表现为X连锁。HGPRT基因长约34kb,而成熟的mRNA的长度只有1.6kb,可见DNA分子内含有大量的内含子。HGPRT的突变基因及表达产物(mRNA,蛋白质)已被分离、研究,和血红蛋白一样有许多突变类型,如有的109位丝氨酸被亮氨酸代替,有的103位丝氨酸被精氨酸代替,还有报告基因发生缺失、重排的严重发病患者。1983年Wilson等发现一例DNA上的TagI限制性内切酶的切点发生突变,使在正常情况下可切出2.0kb片段的电泳区带,由于切点突变而失去酶切作用,变成4.0kb的区带出现。在蛋白质水平进一步研究发现50位的精氨酸被甘氨酸取代。
三、诊断分子生物学在生化遗传学实验室的发展前景
在我国医学检验领域,60年代蛋白质研究的兴起,70年代酶学研究的形成,80年代单克隆抗体的开发为临床生化检验建立灵敏、特异的检测方法奠定了基础。许多检验指标在临床诊断中起着非常重要的作用。
遗传性疾病的发病基础是核酸分子结构变异与核酸的表达产物蛋白质、酶分子结构改变所致。前者可通过染色体形态观察加以分析,后者可通过电泳方法或酶活性测定加以分析。这些方法与免疫方法结合,使检测方法特异性更好,灵敏度更高。这些已建立的方法在遗传性疾病的诊断,蛋白质、酶的多态分析中起重要作用,目前仍为实验室所广泛采用。核酸研究的发展与诊断分子生物学的形成,使生化遗传学研究进入到核酸分子水平,给先天性遗传病治疗带来了希望。
90年代随我国对外开放力度不断加大,对外交流日益加强,诊断分子生物学在我国形成,发展异常迅速,不断地向临床各学科各领域渗透。过去认为遗传性疾病无法根治,但基因研究在临床研究与应用的迅速发展,基因治疗的初步形成,已有治疗成功的先例。基因治疗的发展必然给我们基因诊断提出新的课题、新的要求。诊断分子生物学的发展速度,基因诊断的水平必将影响基因治疗在临床开展的进程。
近年来,分子生物学的新技术、研究成果和学科生长点频频出现,不仅仅在解决人类的疾病诊断、治疗方面成绩显著,在解决粮食、环境等棘手问题也带来新的希望。分子生物学研究领域很多,简而言之主要涉及两大方面内容:基因结构与基因调控。基因结构研究就是揭示人基因组的序列分析,该项工作耗资巨大,取得很多成果,但人基因组非常复杂,有待新的快速测定方法开发,加快研究速度。基因调控研究更为复杂,参与基因调控的元件、因子之多,令人眼花缭乱。分子生物学技术及研究手段也有多种多样,但目前在临床应用得较多的主要有杂交技术和扩增技术。