第四章 血浆蛋白及其代谢紊乱
血浆脂类包括游离胆固醇(freecholesterol,FC)、胆固醇酯(cholesterolester,CE)、磷脂(phospholipid,PL)、甘油三酯(triacylglycerol/triglyceride,TG)、糖酯、游离脂肪酸(freefattyacid,FFA)等。血浆中最多的脂质有胆固醇(总胆固醇totalcholesterol,TC)、PL和LG,血浆脂质总量为4.0-7.0g/L。
血浆脂类简称血脂,其含量与全身相比只占其小部分,然而其代谢却非常活跃。肠道吸收的外源性食物酯类、肝合成的内源性脂类及脂肪组织贮存的脂肪动员都必须先经血液再到其他组织,因此,血脂水平可反映全身脂类代谢的状态。由于血脂的不断降解和重新合成在正常地进行,并保持动态平衡,血脂含量的变动也就稳定在一定的范围内。测定血浆脂类可及时地反映体内脂类代谢状况。就测定方法而言,从历史上看,测定血浆胆固醇是最古老的先行方法,其后陆续进行PL、TG和FFA的定量测定,尔后又增加脂蛋白及其载脂蛋白的测定,这些项目,已是目前临床上用于了解人体脂类代谢状况的系列指标。
第一节 血浆脂蛋白
一、血浆脂蛋白的分类
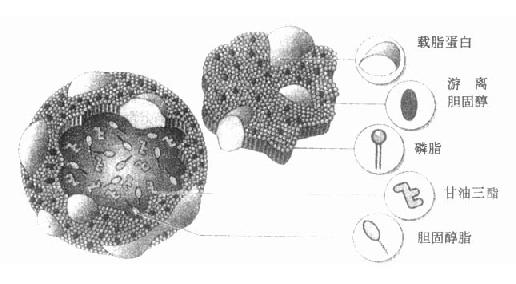
脂蛋白属于一类物质,因结构及组成的差异,有多种形式存在,尽管如此,仍有许多共同之处,一般都是以不溶于水的TG和CE为核心,表面覆盖有少量蛋白质和极性的PL、FFA,它们的亲水基因暴露在表面突入周围水相,从而使脂蛋白颗粒能稳定地分散在水相血浆中,如图4-1所示。
图4-1 脂蛋白结构图
血浆脂蛋白的分类方法主要有电泳法和超速离心法
(一)超速离心法
超速离心法是根据各种脂蛋白在一定密度的介质中进行离心时,因漂浮速率不同而进行分离的方法。脂蛋白中有两种比重不同的蛋白质和脂质,蛋白质含量高者,比重大;相反脂类含量高者,比重小。从低到高调整介质密度后超速离心,可依次将不同密度的脂蛋白分开。通常可将血浆脂蛋白分为乳糜微粒(chylomicron,CM)、极低密度脂蛋白(verylowdensitylipoprotein,VLDL)、低密度脂蛋白(lowdensitylipoprotein,LDL)和高密度脂蛋白(highdensitylipoprotein,HDL)等四大类。
(二)电泳法
由于血浆脂蛋白表面电荷量大小不同,在电场中,其迁移速率也不同,从而将血浆脂蛋白分为乳糜微粒、β-脂蛋白、前β-脂蛋白和α-脂蛋白等四种。α-脂蛋白中蛋白质含量最高,在电场作用下,电荷量大,分子量小,电泳速度最快,电泳在相当于α1球蛋白的位置。CM的蛋白质含量很低,98%是不带电荷的脂类,特别是甘油三酯含量最高。在电场中几乎不移动,所以停留在原点。为了取样方便,多以血清代替血浆。正常人空腹血清在一般电泳谱上无乳糜微粒。电泳分类法的脂蛋白种类与超速离心法的脂蛋白分类相应关系如图4-2所示。
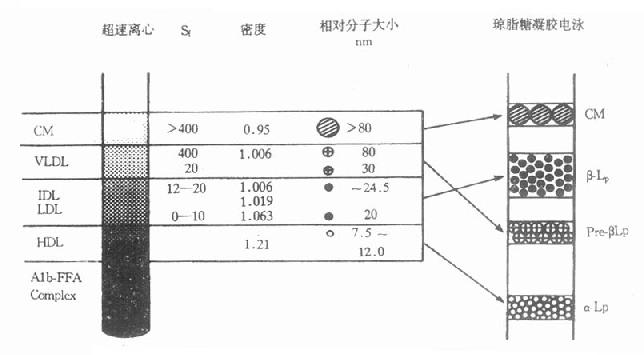
图4-2 超速离心法与电泳法分离血浆脂蛋白的相应名称
二、脂蛋白组成与结构
一般认为血浆脂蛋白都具有类似的结构,呈球状,在颗粒表面是极性分子,如蛋白质,磷脂,故具有亲水性;非极性分子如甘油三酯、胆固醇酯则藏于其内部。磷脂的极性部分可与蛋白质结合,非极性部分可与其它脂类结合,作为连接蛋白质和脂类的桥梁,使非水溶性的脂类固系在脂蛋白中。磷脂和胆固醇对维系脂蛋白的构型均具有重要作用。
1.乳糜微粒CM颗粒最大,约为500nm大小,脂类含量高达98%,蛋白质含量少于2%,因此密度极低。CM由小肠粘膜细胞在吸收食物脂类(主要是甘油三酯)时合成,经乳糜导管,胸导管到血液。主要功能为运输外源性甘油三酯。
2.极低密度脂蛋白VLDL中TG主要在肝脏利用脂肪酸和葡萄糖合成。若食物摄取过量糖或体内脂肪动用过多,均可导致血VLDL增高。VLDL中脂类占85%-90%,其中TG占55%,其密度也很低。VLDL是运输内源性TG的主要形式。
3.低密度脂蛋白LDL的结构大致可分为三层:内层,占15%的蛋白质构成核心,被一圈磷脂分子包围;中层,非极性脂类居中,并插入内外层,与非极性部分结合;外层,85%的蛋白质构成框架,磷脂的非极性部分镶嵌在框架中,其极性部分与水溶性的蛋白质等亲水基团突入周围水相,使其脂蛋白稳定地分散于水溶液中;游离胆固醇分布于三层之中。
4.高密度脂蛋白HDL是一组不均一的脂蛋白,经超速离心和等电聚焦电泳,可把HDL分成若干亚族。各亚族具有不同的密度,颗粒大小及分子量不尽相同,脂质和载脂蛋白比例不同,经X射线衍射研究证实为三维形态结构。现有资料提示,HDL是对称的准球形颗粒,具有一低电子密度的核心的外壳。低电子密度的中心由非极性脂质所占据,高电子密度是部分由磷脂极性头和蛋白质组成的颗粒外壳。经园二色分析证实,HDL的蛋白部分有2/3是α-螺旋结构,其余为无规则结构。带电荷的极性氨基酸残基构成α-螺旋的极性面,而疏水侧链则占据另一面。氨基酸按顺序排列在螺旋区域形成两性结构。目前认为,HDL的结构是α螺旋区平行于脂蛋白颗粒表面,非极性氨基酸残基伸展到颗粒的非极性核心区域;磷脂的脂肪酰链则垂直于脂蛋白颗粒表面的螺旋形载脂蛋白;胆固醇酯深埋在HDL颗粒的亲脂核心内;而游离的胆固醇可能与颗粒表面在磷脂极性头和载脂蛋白结合。
HDL主要由肝合成,小肠也可合成。HDL按密度大小又可分为HDL1、HDL2和HDL3。HDL1又称为HDLc,仅在摄取高胆固醇膳食后才在血中出现,健康人血浆中主要含HDL2和HDL3。HDL主要是将胆固醇从肝外组织转运到肝进行代谢。
5.脂蛋白(a)Berg于1963年在血浆脂蛋白电泳时发现β-脂蛋白部分有一种新的抗原成分,并与LDL结合,将此抗原成分命名为脂蛋白(a)[lipoprotein(a),LP(a)]。其后证实,LP(a)核心部分由甘油三酯、磷脂、胆固醇、胆固醇酯等脂质和载脂蛋白B100组成,结构类似LDL,并含有LDL中没有的载脂蛋白(a)[apolipoprotein(a),Apo(a)]。Apo(a)与纤溶酸原具有高度同源性,在纤溶系统多个环节发挥作用,从而影响动脉粥样硬化性疾病的发生和发展。有足够证据表明,Lp(a)是动脉粥样硬化性疾病的一项独立危险因子。Lp(a)含有两类载脂蛋白,即ApoB100和Apo(a),两者通过1至2个二硫键共价相连,若用还原剂巯基乙醇处理Lp(a)时,Apo(a)可从Lp(a)的分子上脱落下来,成为不含脂质的一类糖蛋白。剩下不含Apo(a)仅含ApoB100的颗粒,称为Lp(a-)。