�ı�
�� ��2
֢ ��4
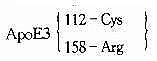
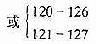
�����ڡ���֬������Ķ�̬��
������������������У������DNA���������ݱ䡣�����ݱ�����ڰ�������������������Ϊ�����ԡ��仯���������ٴ������ķ��������ǣ�����İ�����������Լ��������ĵ�������DNA���иı�ʱ�з��������µ����ʵĺϳ����ı������ܵ��쳣����֬���ṹ������������DNA���е�ͻ�������¸�֬Ѫ֢���緢��������Ӳ���Ѵ���������ҽѧ���ס��µ���֬������Ķ�̬�Բ��ϵر����֡�
���������֬��������Ŵ�����ͨ��ʹ�ô�����Ѫ��ϸ������ȡ��DNAΪ��ʼ���ϣ�ͨ���ۺ�ø����Ӧ��������DNA�IJ������ú���ñ����ݶ�������Ӿ��denaturing gradient gel electrophoresis, DGGE����ʾ�ɻ�����밷������DAN����ͻ�����µĵ���DNA����Ķ�̬�ԣ���ʹ�õ�λ��������ĹѺ�����̽���ӽ�����allele-specific oligonuc leotide hybridization��ASO��̽��ͻ��ĵ�λ����ⶨ������Ƭ�γ��ȶ�̬�ԣ�restriction fragment length polymorphism, RFLP����������DNA���е�ͻ�䣻��ֱ�Ӷ�ij�����Ƭ�ν���DNA���вⶨ������������Ӧ��DNA���бȽϣ�����з�ͻ�䷢�����⼸�ַ���Ϊ�ϳ�ʹ�õķ������й���Щ��������ϸ���ܿɲμ������ʮ���µ����ﻯѧ���鼼��ƪ��������Ҫ����ApoB��ApoE,Apo��a����ApoA�����Ķ�̬�ԡ���������֬������Ķ�̬�Խ��Ծ����ķ�ʽ������
����һ��ApoB������Ŵ��������̬��
����ApoB������Ŵ��������̬�Է����¼������ͽ��н��ܣ���RFLP���ڵ���Ѫ�����̴�ˮƽ���͵Ļ���ͻ�䣻�۵���Ѫ�����̴����ߵĻ���ͻ�䣻���������͵Ķ�̬�ԡ�
����1��RFLP
�����ݱ�������֬����B�����������Ƭ�γ��ȶ�̬������375�֡�����������Щ��̬�Բ��Ǵ��ڣ�����DNA���вⶨ����4-3�о���10��RFLP��������ЩRFLP��ͻ�������ApoB�����������������������ں�������������������
��4-3 ApoB����������Ƭ�γ��ȶ�̬�Ծ���
| ������������ø | ø��λ�� | ͻ���λ�������Ƶ�� | ø�в�λ���ڻ�ȱʧ |
| Ava�� | ���1����������4Kb | 0.20 | ȱʧ |
| Msp�� | ���������ڣ�-265bp | 0.21 | ȱʧ |
| ApaL�� | ��4�����ӣ�cDNA��417bp | 0.36 | ȱʧ |
| Hinc�� | ��4�ں����ڣ����3������3'ĩ��3334bp | 0.12 | ���� |
| Pvu�� | ��4�ں����ڣ���5������5'ĩ��4523bp | 0.08 | ���� |
| Alu�� | ��14�������ڣ�cDNA��1981bp | 0.48 | ȱʧ |
| Bal�� | ��20�ں����ڣ����21������5'ĩ��146bp | 0.50 | �� |
| Xba�� | ��26�������ڣ�cDNA��7674bp | 0.40-0.50 | ���� |
| Msp�� | ��26�������ڣ�cDNA��11040bp | 0.12 | ȱʧ |
| EcoR�� | ��26�������ڣ�cDNA��12670bp | 0.20 | ȱʧ |
������Դ�ԣ�Young S G. Circulation 82:1583,1990��
������4-3���з������������ڵ�ͻ�䣬��Xba��������Ƭ�γ��ȶ�̬���⣬��������°�����ı䡣���кܶ��о����������о�������ApoB����PFLP��ߵ���Ѫ֢���߸�������Ѫ֢Ӳ������ع�ϵ�������������ŷ�����ع�ϵ�ı������Ǻܶ࣬���еĽ���ì�ܡ�����Youngָ�����������е��о���ʹ�õ�������̫С���е��о��������ѡ�Ϲ淶���е��о����ڲ�ͬ�����壬������������������졣����Ȼ�����Ժ������о��У�������ѡ�ô���������������ѡ������飬���Dz�ͬ����������Ŵ��������ɶ�ȷ֤ijЩApoE�����RFLP��֬���״�л���ҵ���������Լ��緢��������Ӳ���Ĺ�����Ϊ��Ҫ��
����2������Ѫ�����̴�ˮƽ���͵�ApoB����ͻ��
������������ʵ���ҵ�Ŭ����һЩApoB���������ͻ�������ͻ�䵼�¼������ܶ�֬����Ѫ֢���Ѽ���������4-4����
��4-4 ApoB����ͻ�䵼�¶�ApoB�ĺϳ�
| ApoB�ij��� | DNAˮƽ�ϵ�ͻ�� | ��ApoB��Ѫ��֬�����еķֲ� |
| B-2 | �ڵ�5�ں������У�G����T | �� |
| B-9 | T���C��ʹ������411�������Ӻ���ֹ | �� |
| B-25 | �ڵ�21����������694bpȱʧ | �� |
| B-27.6 | �ڵ�24�ں����ڣ����Ӳ�λͻ�� | HDL��LDL |
| B-29 | �ڵ�4152�������ϣ�T���C | �� |
| B-31 | cDNA������4480ȱʧ | HDL���ܶȴ���1.2g/cm3���� |
| B-32 | ��������1450�ϣ�T���C | HDL��LDL |
| B-32.5 | ��4631�������ϣ�G���T | HDL���ܶȴ���1.21g/cm3���� |
| B-37 | cDNA������5391��5394��ȱʧ | VLDL��LDL��HDL |
| B-39 | cDNA������5591ȱʧ | VLDL��LDL |
| B-40 | cDNA������5693��5694��ȱʧ | VLDL��LDL��HDL |
| B-46 | ��cDNA������6381�ϣ�T���C | VLDL��LDL��HDL |
| B-49.6 | ��cDNA������6963�ϣ�T���C | VLDL��LDL��HDL |
| B-52.8 | cDNA������7295ȱʧ | VLDL��HDL |
| B-54.8 | cDNA�˸���7539ȱʧ�� cDNA������7655�ϣ�T���C |
VLDL��HDL |
| B-61 | �ں�����8525������37bpȱʧ | VLDL��HDL |
| B-67 | cDNA������9327ȱʧ | LDL��VLDL |
| B-74.7 | cDNA������10366ȱʧ | LDL��VLDL |
| B-82 | �ں�����11411�ϣ�A���C | VLDL��HDL |
| B-86 | cDNA������11840ȱʧ | VLDL��HDL |
| B-87 | cDNA������12032ȱʧ | VLDL��HDL |
| B-89 | cDNA������12309ȱʧ | VLDL��HDL |
����(Դ�ԣ�Kane ,J.P., and Havel, R.J.In:The metalbolic and molecular bases of inherited disease, edited by Scriver, C.R.et al, Vol ��,p1868,1995,McGraw �CHill,Inc.,New York, U.S.A)
���������Ե͵��ܶ�֬����Ѫ֢�Ļ��ߴ��Ϊ�Ӻ��ӣ�������Я��һ��������ApoB��λ�����һ�������ApoB��λ���������Լ������Ӻ��ӻ����ټ�����ApoB�ϳɵĻ���Ѫ����ApoB��Ũ�Ƚϵͣ���Ϊ������ApoB��λ���������ApoB-100Ũ�ȵ�2%��10%���������δ��ȫ���ˡ����˷��֣�����ApoB87��ApoB89��LDL�Ⱥ�ApoB-100��LDL�����ڱ�LDL������ȡ���������Ʋ����Ӻ��ӻ����У���һ������ApoB-100��λ��������ApoB-100����������ApoE��λ��������ApoE��ɵ�֬��������������֬�������ຬ��1����ApoE����ǰ�����ڱ�LDL��ȡ�������Щ�������ڽ���һ���ֶ�ApoB�ϳɻ������ڵ�Ũ��Ѫ��ApoB����Ũ��Ѫ�����̴��Ļ����еȽ�һ��Ŭ���������뵽���ǣ������Ե͵��ܶ�֬����Ѫ֢����Ѫ����LDL���̴��ͣ��������IJ�����������Ӳ����Σ����Ҳ���͡�����һЩ����֤ʵ����Щ���߲�������������Ӳ����������������������Ϊ���Թ߳�ʳ�ø߱���֬�����ߵ��̴���ʳ�ı�����Ⱥ��Я���ϳɶ�ApoB��ͻ�����ǵ��������������ڽ�����
����3������Ѫ�����̴����ߵĻ���ͻ�䡣
����Brown��Goldstein�ľ����о�����LDL��������ͻ�䣬����LDL����ϰ���LDL��Ѫ��������������緢��������Ӳ�������Ǽ����Ըߵ��̴�Ѫ֢����Ҫ������Щ���ߵ�Ѫ���ܵ��̴�Ũ�ȸ���7.76mmol/L��Ȼ�����൱һ�����˵�Ѫ�����̴�Ũ����6.47��7.76mmol/L֮�䣬�����������Ϊ���ж����ߡ���������������һֱ�Ʋ�ApoB�����Ŵ�����ɺϳ���ȱ���ApoB�����ʣ�������LDL���������½�������LDL����ϰ���LDL��Ѫ��������������Ʋ���1989��õ���֤ʵ��Soria���˷����˼�����ApoB100ȱ����Щ��������Apob cDNA��10708��������λ���ϣ�Gת������A������ApoB100�ĵ�3500�������ᾫ����任���˹Ȱ���������һͻ��ʹ��LDL��LDL����Ľ�����½��������ߵ��̴�Ѫ֢���ݹ��ƣ�����ͻ������Ⱥ�еķ�������LDL�����ͻ���ʽӽ�����1/500��
������һ������ApoB100ȱ������ApoB�����10800������λ���ϵ�Cת����T���¡���һͻ��ApoB�ڵ�3531�������ᾫ�������˰��װ��ᡣ8�����ߵ�ƽ��Ѫ��LDL���̴�ˮƽ��6.78mmol/L����8��δ��ApoB����ͻ�������Ϊ3.88mmol/L������LDL��LDL�����������Ϊ�����������39%������C��T��ת��������һ���µ�����������ø�иλ��NsiI��,�⽫�������ڴ�Χ��Ⱥ�ڼ������ͻ�䷢���ʡ�
����4���������͵Ķ�̬��
������ApoB����3'ĩ��500������������������һDNAƬ�ι��ɿɱ䶯��Ŀ�İ����ظ���variable number of tandem repeats. VNTR���ѷ���ApoB����3'ĩ������VNTR���µĶ�̬�����ļ������ķ����й�����
����ApoB�ź�����27����������ɡ��ڸ���������Ⱥ�з��������ֶ�̬�ԣ�һ��Ϊ�����λ�����ͣ�����27���������һ��Ϊȱʧ��λ�����ͣ�����24�������ᡣVisvikis ���˷���ȱʧ��λ���������Ѫ��ApoB ˮƽ��LDL���̴�ˮƽ��LP��a��ˮƽ�Լ��ܵ��̴�ˮƽ�йء����Kammerer������34�ҹ�686������ī�������з��ֱ���ApoB�ź��ĵ����ֵ�λ����SP-24��SP-27��SP-29�����Ƿֱ����24��27��29�ġ�Я��SP-24��λ����Ĵ�������Ѫ��ApoB��LDL���̴���Ũ����������SP-27�����ӡ�SP-29�Ӻ��ӵ�ApoB��LDL���̴�ˮƽ��SP-24�����ӻ��ߡ�
��������ApoE����Ķ�̬��
����ApoE�ڵ��̴������������л��������Ҫ���á���������Ⱥ�У�10%���ϵĸ����Ѫ�����̴�ˮƽ������ɹ龿��ApoE����Ķ�̬�ԡ���Ҫ��ApoE��λ������ApoE2��ApoE3��ApoE4��ApoE3����Ⱥ�еij�������ߣ�����Ϊ�ϳ�����ApoE����ApoE2�����֬����Ѫ֢��أ���ApoE4�������Գմ�����Alzheimer's disease�������еij����ʽϸߡ����һ���ֿ��������ǵ���Ұ����ApoE������֬���л��������Ҫ�������������Ĵ�л������Ҳ������Ҫ��ɫ�����б�����ApoE������֯���˵��������ߵ����Լ�����ϸ��������ֻ�����������ʱ�����ƣ������µ�ApoE���ܼ��µ�ApoE�����̬�Իᱻ���֡�
������4-5�о���һЩ�Ѽ�����ApoE������Ŵ����졣����ͬʱ�г��������Ӽ������İ�����ĸı䡢�ض�ApoE������������ȱ����������Ǹ����塢����Ƶ�ʡ���صĸ�֬Ѫ֢���Լ��Ŵ���ʽ��
��4-5 ApoE��̬��
| ��̬������1 | �������롡�� | ������ �ı� |
������
�� ��2 |
���屳�� | �ֲ���3 | ��֬Ѫ
֢ ��4 |
�Ŵ���ʽ | |
| ������ApoE������ | ||||||||
|
- | - | - | �������� | 41.9��91.1% | - | �� | |
| ApoE4 | 112 | TGC��CGC | Cys��Arg | �������� | 6.4%��36.8% | N/HC | �� | |
| ApoE2 | 158 | CGC��TGC | Arg��Cys | ������ | 0.0%��14.5% | N/FD | ���� | |
| ��FD��صĶ�̬�� | ||||||||
| ApoE0 | ��3�ں��� | A ��G,Gȱ | �ϳ����ֶ��� | ������ | 14 | FD | ���� | |
| 3592bp-60 | ʧ����ͻ�� | �ϳ�60�� | �¹��� | 3 | FD | ���� | ||
| ApoE4-�ѳ� | 13 | GAG ��AAG | Glu ��Lys | ���������� | 7 | FD | ���� | |
| 145 | CGT ��TGT | Arg ��Cys | ���������� | �� | �� | ���� | ||
| ApoE3-Leiden | 112 | TGC ��CGC | Cys ��Arg | ������ | >40 | FD | ���� | |
|
21bp�ظ� | 7�������Ỽ���ظ� | �� | �� | �� | �� | ||
| ApoE1 | 127 | GGC��GAC | Gly��Asp | ������ | >25 | FD | δ֪ | |
| 158 | CGC��TGC | Arg��Cys | �� | �� | �� | �� | ||
| ApoE2-Christchurch | 136 | CGC��AGC | Arg��Ser | δ���� | 3 | FD | δ֪ | |
| ApoE3 | 112 | TGC��CGC | Cys��Arg | ���������� | 6 | FD | ���� | |
| 142 | CGC��TGC | Arg��Cys | �� | �� | �� | �� | ||
| ApoE3--kochi | 145 | CGT��TGT | Arg��His | �ձ��� | 4 | FD | δ֪ | |
| ApoE2 | 145 | CGT��TGT | Arg��His | δ���� | 3 | FD | δ֪ | |
| ApoE1- Harrisburg | 146 | AAG��GAG | Lys��Glu | �ձ��˸��� �� | 8 | FD | δ֪ | |
| ApoE2 | 146 | AAG��GAG | Lys��Gln | ������������ | >40 | FD | ���� | |
| ApoE3��ʢ�� | 210 | TGG��TAG | Trp����ֹ���� | �������� | 3 | FD | ���� | |
| ApoE2 Fukuoka | 224 | CGG��CAG | Arg��Gln | �ձ��� | 1 | FD | δ֪ | |
| ���FD��֬Ѫ֢��صĶ�̬�� | ||||||||
| ApoE5 | 3 | GAG��AAG | Glu��Lys | �ձ��� | 0.5% | HC | �� | |
| 13 | GAG��AAG | Glu��Lys | �������� | 6 | HC/HTG | �� | ||
| ApoE2-Dunedin | 228 | CGC��TGC | Arg��Cys | ��� | 2 | HTG | �� | |
| ApoE2 | 236 | GTG��GAG | Val��Glu | ������ | 8 | HTG | �� | |
| ApoE7-Suita | 244 | GAG��AAG | Glu��Lys | �ձ��� | 0.5%��0.8% | HC/HTG | �� | |
| 245 | GAG��AAG | Glu��Cys | �� | �� | �� | �� | ||
| ApoE3 | 112 | TGC��CGC | Cys��Arg | ������ | 3 | HTG | �� | |
| 251 | CGC��GGC | Arg��Gly | �� | �� | �� | �� | ||
| ApoE1 | 158 | CGC��TGC | Arg��Cys | ������ | 2 | HC | �� | |
| 252 | CTG��GAG | Leu��Glu | �� | �� | �� | �� | ||
| ���֬Ѫ֢�صĶ�̬�� | ||||||||
| ApoE4-+-Ferburg | 28 | CTG��CCG | Leu��Pro | �¹������� | 0.4%��0.9% | N | �� | |
| 112 | TGC��CGC | Cys��Arg | �� | �� | �� | �� | ||
| ApoE4 | 112 | TGC��CGC | Cys��Arg | ������ | 5 | N | �� | |
| 274 | CGC��CAC | Arg��His | �� | �� | �� | �� | ||
| ApoE4+ | 296 | AGC��CGC | Ser��Arg | ������ | 5 | N | �� | |
| ApoE5 | 84 | CCG��CGG | Pro��Arg | �������� | 0.2% | N | �� | |
| 112 | TGC��CGC | Cys��Arg | �� | �� | �� | �� | ||
| ApoE3- Freibaug | 42 | ACA��GCA | Thr��Ala | �¹������� | 1 | N | �� | |
| ApoE3 | 99 | GCG��ACG | Ala��Thr | ������ | 1 | N | �� | |
| 152 | GCC��CCC | Ara��Pro | �� | �� | �� | �� | ||
| ApoE2 | 134 | CGG��CAG | Arg��Gln | ������ | 5 | N | �� | |
����1�����ѳǣ�Philadelphia���뻪ʢ�٣�Washington���������⣬�����ĵ�����δ���������
����2����������ţ�Cys-���װ��Arg-�����Glu-�Ȱ��Lys-�����Gly-�ʰ��Asp-���Ŷ����Ser-˿���His-�鰱�Gln-�Ȱ�������Trp-ɫ���Val-�Ӱ��Leu-�����Thr-�հ��Ala-�����Pro-�����
����3���ֲ��ʣ��Ӱٷֺŵķֲ����Ǹ��ݼ��1000�����ϵĽ������ģ�δ�Ӱٷֺŵ���ֵ���������Я��ApoE��̬�ͣ��������������Ӻ��ӣ���������
����4����д��N��Normal,������HC��hypercholesterolemia ���ߵ��̴�Ѫ֢����FD��familial dysbetalipoproteinemia,�������쳣���ܶ�������Ѫ֢��������֬����Ѫ֢����
������Դ�ԣ�Knijff et al. Hum. Mutation 4:181,1994��
��������Apo��a������Ķ�̬��
����Ѫ��Lp��a����Ũ����Apo��a�������ʵĴ�С�ɷ��ȣ���Apo��a���ķ�����������Lp��a����Ũ�����ͣ�Apo��a���ķ�������С����Lp��a����Ũ�����ߡ�Apo��a���ķ�������400��800KDa֮�䡣����Apo��a���ھ۱�ϩ��������ʱ�����ApoB100��������Ϊ520KDa����Ǩ���ʣ�Utermann������1987�꽫Apo��a������Ϊ���ֶ�̬�ͣ�F��ApoB100Ǩ�ƿ죻B��ApoB100Ǩ������ȣ�S1��S2��S3��S4��ApoB100Ǩ����������֤ʵ��Apo��a�������ʵĶ�̬������Apo��a��������Kringle-4����Ŀ�仯������ġ�Kring-4����Ƭ�κ��м���ϡ�ٵ�������ø�в�λ��NotI,SfiI.KspI,Sval,KpnI������Ϊ����ɸѡApo��a������Ķ�̬���ṩ�˰�������4-5�о��ڰµ�������Tyrolean��������Ⱥ����KpnI����������õ�RFLP��1-4Kringe����Ŀ��Apo��a�������ʵĴ�С�Լ�Ѫ��Lp��a��Ũ�ȵĹ�ϵ��
�� ��4-6 Apo��a��Kpni Ƭ����Apo��a�������ʶ�̬�͡�Kringle-4��������Ŀ�Լ�Ѫ��Lp��a��Ũ��֮��ϵ
| �� | ���ַ�����(KDa) | KpnIƬ��(Kb) | Kringle-4��Ŀ | ��λ���� ��Ŀ | ƽ��Lp(a)Ũ��(mg%) | KpnIƬ��Ƶ��(%) |
| F | ��450 | 37-49 | 11-13 | 1-3 | * | ��0.2 |
| B | ��500 | 55-66 | 14-16 | 4-6 | 61.7 | 1.2 |
| S1 | ��550 | 72-82 | 17-19 | 7-9 | 34.4 | 3.8 |
| S2 | ��600 | 88-99 | 20-22 | 10-12 | 24.5 | 11.2 |
| S3 | ��650 | 105-116 | 23-25 | 13-15 | 10.2 | 13.7 |
| S4 | ��700 | 121-210 | 26-42 | 16-32 | ��5.7 | 70.1 |
����*:������̫С,����ȱ�硣(Դ�ԣ�Utermann,G.In:The metabolic and molecular bases of inherited disease.7th ed, Vol.��,eds.Scriver,C.R.et al.p1897,McGraw-Hill.Inc.,New York,1995)
�����ɱ�4-6�ɿ�����Ѫ��Lp��a����Ũ������KpnƬ�εĴ�С�ɷ��ȡ���ΪѪ��Lp��a����Apo��a����̬�͵Ĵ�С�ɷ��ȴӻ���DNAˮƽ�ṩ��ֱ�ӵ�֤�ݡ�
����Scana������Kringle-4��37���ظ�����KIV-37���ֳ�ΪKringle��-10������������ͻ�䣺һ��ͻ�䵼�µ�72�������ᱻ������ȡ������һ��ͻ�䵼�µ�66���������հ��ᱻ������ȡ����ǰ����ʹ��Lp��a���������������ϣ���Ѫ��Lp��a��Ũ�Ƚ��͡������ƺ���Lp��a��Ũ���ء�Mooser���˱����ھ��1��������5����1.3Kb�����ڣ���5��������[��TTT-TA��n]���ɵ�VNTR����Ѫ��Lp��a��Ũ����ء����⡣���б���Apo��a������5���˲�����DNA���еĶ�̬�Կ���Ѫ��Lp��a����Ũ�ȡ�
�����ġ�ApoA�����Ķ�̬��
����ApoA��ΪHDL����Ҫ�ṹ���ס�ApoAI����ͻ��ɵ����쳣��ApoA���ʵĺϳɡ���Щ�쳣��ApoA��Ӱ��HDL�Ĵ�л���ѱ������谭ApoA��ϳɵĻ���ͻ�������š�ȱʧ������ͻ��ȷ�ʽ����Щͻ��Ĵ����ӻ��߱��ֳ�����HDL���̴�ˮƽ���緢���IJ���Ŀǰ�ѷ���������20�ֲ�ͬ��ApoAI�ṹ����ĵ�ͻ�䵼�°�����ת��������4-7������������ͻ�䣨������173�����װ��������165�������ᣩ���HDL���̴�ˮƽ��أ������������緢���IJ���
����Talmud���˷���ApoA����������������ھ�ת¼��ʼ��λ��75�������-75bp��A�û�G��ͻ�����ApoAI��HDLˮƽ��ء�����ͻ���ĵ�λ�����ΪA��λ������G�ij�ΪG��λ����Saha�ȱ���A��λ�������¼��»����г��ֵ�Ƶ�ʣ�0.27�����ڸ�����(0.12)��Wang���˷��֣�ApoA������1�ں���5'ĩ�ˣ���83���Cת����T���84���Gת����A����ʹMspIø�е�ɥʧ������ͻ�����HDL���̴���أ���������-75���A�û�Gͻ�����HDL���̴�����ػ�ǿ��
��4��7������ApoAI�����ͻ�䵼�°�����ı�
| ������ı�1 | ������2 | �������ͻ�� |
| Pro3��Arg | CCC | C��G |
| Pro3��His | CCC | C��A |
| Pro4��Arg | CCC | C��G |
| Arg10��Leu | CGA | G��T |
| Gly26��Arg | CGGC | G��C |
| Asp89��Glu | GATC | T��G,T��A |
| Asp103��Asn | CGAC | G��A |
| Lys107��0 | �� | �� |
| Lys107��Met | AAG | A��T |
| Glu136��Lys | CGAG | G��A |
| Glu139��Gly | GAG | A��G |
| Pro143��Arg | CCA | C��G |
| Glu147��Val | GAG | A��T |
| Ala158��Glu | GCG | C��G |
| Pro165��Arg | CCC | C��G |
| Glu169��Gln | CGAG | G��C |
| Arg173��Cys | GCGC | C��T |
| Arg177��His | CGC | G��A |
| Glu198��Lys | CGAG | G��A |
| Asp213��Gly | GAC | A��G |
����1����������ſɲμ���4-5��ע����1�Ű��������Ͻǵ���ֵ��ʾ����ApoA��һ���ṹ�ϵ�λ�ã�2���ӵ��ߵĺ�����Ϊ��ͻ�䲿λ����Դ�ԣ�Eckardstein et al. J.Biol Chem.265:8612,1990��
�����塢������֬������Ķ�̬��
���������Ͻ��ܵ�ApoB��ApoE��Apo��a���Լ�ApoAI�Ļ���һ����������֬���Ļ���Ҳ���ڶ�̬�ԣ����¼�Ҫ���о�һЩ���ӡ�
����1��ApoA��-C��-A������������Ƭ�γ��ȶ�̬��
������ApoA��-C��-A��������ڣ�ʹ��Xmn��Apa��Msp��Pst��Sst��Xba��Taq���Pvu������������ø�Ѽ���10����RFLP��������Щ��̬�Դ���ɱ��백���������DNA����ͻ������,������ЩRFLP��Я���������ӹ��IJ���Σ����ء�����������������øPst����������Ⱥ���Ѽ���DNA����ApoA��cDNA̽��ӡ��DNA�ɲ��3.3Kb��DNAƬ�Σ���ΪP2��λ������2.2Kb��DNAƬ�Σ���ΪP1��λ����P1��λ������������Ⱥ�г����ʽϸߡ�P2��λ�����ڵ�HDL���̴������еij����ʸ��ڶ������6��������Ӣ���������ı�����֤ʵ���Ӻ���P1P2Я����Ѫ��ApoA����HDL���̴���ˮƽ�����ص���P1P2������Я���ߡ�����ʾ��ApoA��-C��-A��������У�Pst��ø�е�Ķ�̬�ԣ�P2��λ�����뽵��Ѫ��ApoA���HDL��ˮƽ�Լ��շ����IJ��йء������Dallinga-Thie����������֬���о���־�ϱ�����������������øXmn����Msp����ApoA��-C��-A��������ڼ�����ϡ�е�λ�������Ѫ�����̴�������������ApoB�Լ�LDL���̴�ˮƽ��������Щ�����ʾ��ApoA��-C��-A������Բ������Ѫ�����̴������������Ĵ�л��
����2��ApoE��-C��-C�����������Ƭ�γ��ȶ�̬��
����λ�ڵ�19��Ⱦɫ���ϵ�ApoE��-C��-C������Ҳ�ж�̬�ԵĴ��ڡ�������RFLP������������ø������Hpa��Taq��Bgl��Dra��Nco���Լ�Bgl��Klasen���˱�����Hpa��������Ƭ�γ��ȵĶ�̬����������֬Ѫ֢�йء�Ȼ��Hpa��������Ƭ�γ��ȵĶ�̬����E2/E2�������ؽӹ�������˵��Hpa���RFLP�Dz�ͬ��ApoE�����һ�ֶ������Ŵ����ӣ���ͻ�����δ������
����3��ApoA������ͻ��
����Deeb�ȱ�������һ���ձ������м����ļ�����ApoA��ȱ��֢����״������Ӱ��ʾ��Խ���δ��Ѫ�ܼ���������������������ⲻ��Ѫ��ApoA��ˮƽ������HDL���̴���������Χ��DNA������ʾ������ApoA������ڵ�3�ں��Ӽ��ӹ��岿λ�ĵ�һ�����Gͻ���ΪA���Ӷ��谭���ں��Ӵӳ���ת¼���м������Ĺ��̡��������ǵ���ApoA��ȱ�ݵ�ԭ��
����4��ApoA������Ķ�̬��
����������ApoA����λ�������ApoA���ĵ�360��347�������ᣬ�ֱ�Ϊ�Ȱ��������հ��ᡣ��ApoA������ĵ�360�������ϵ�3�����T�û�G�������ĵ�360������λ���ϵĹȰ���������֮Ϊ�鰱�ᣬ����347�������ϵ�1�����T�û�A���հ���347��˿����ı��켴����������������ApoA�������̬�Զ�֬���л��Ӱ������һ�µĽ��ۡ���һ��ͻ�䣬��������127��ͻ�����Hinc��ø��λ���ɥʧ�����Kamboh���˶����������в���ѱ¹�������з��֣�û��Hinc��ø��λ���λ�����Я����Ѫ����������ˮƽ����Hinc��ø��λ���λ�����Я���ߡ�
����5��ApoC�����Ķ�̬��
������֬����C������ͻ�����������ͱ�������a.����ͻ�䣻b.�������ӵļ��ͻ�䣻c.��2�ں��Ӽ��й��岿λͻ�䣻d.�ɼ�������ȱʧ���µ�����ͻ�䡣��Щͻ�仼�ߵ�Ѫ��ApoC��ˮƽ�������ͻ�ȱʧ�����Ŵ���ʽΪ��Ⱦɫ�������Ŵ���
����6��ApoC�����Ķ�̬��
�����ѷ���������ApoC������̬����߸�������Ѫ֢�йأ�a.3175������C����G��b.3206������T����G��c.3206G��λ����Li���˲���ǰ���ٴ��о���־�ϱ�����ApoC�������������-482��-455�����ͻ���뷢���߸�������Ѫ֢�йء������Ʋ�����ͻ�������������ȵ�������DNA���з�Χ�ڡ�ͻ����ApoC����������Ӱ������ȵ��ص����Ƶ��أ�ApoC���ת¼����ǿ��ApoC��ĺϳ����ӡ����������Ⱥ�з����߸���������һ����Ҫ���ء�
����7��ApoD����Ķ�̬��
����Vijayaraghavan���˼����57�����ֲ���57��������ApoD����Taq��ø��λ��������Ƭ�γ��ȶ�̬�ԣ�2.2��2.7Kb��λ�������Ƿ���2.2Kb��λ�����ڷ������еķֲ������ظ��������ߡ����������2.2Kb��λ������Ӻ����봿���ӣ���ʾ�����Ŵ���ʽ��������Ϊ��ApoD����Taq��������Ƭ�γ��ȶ�̬�Կ���Ϊ�о����ֲ���һ���Ŵ���־��
����8��ApoH����Ķ�̬��
����ApoH�����247������ͻ�䣬��ʹ��247���������Ӱ��ᱻ������ȡ������Ϊ��ͻ��ʹRas��ø��λ��ɥʧ��������������Ƭ�γ��ȶ�̬�Ժ���⡣���Kamboh�ȵ������������в�ѱ¹����������ApoH��λ����ApoH1��ApoH2����ApoH3�ķֲ���Ѫ��֬��ˮƽ�Ĺ�ϵ�����Ƿ���ApoH��λ���������ӵ�������ˮƽ��ء�����Ů�Ӹ߸�������ˮƽ��ء���Ů�Ӹ߸�������ˮƽ��ء�ApoH����Ķ�̬����֬�ʴ�л�Ĺ�ϵ���д��ڸ��㷺�ء��������̽�֡�
����9��ApoJ����Ķ�̬��
����Kamboh��ͬ�±������ڶ����������ˡ����ˡ�����ӡ�ڰ��ˡ���˹��Ħ�ˡ��¼������˼�����������6�����干985�������ߵļ���С����Ƿ�����ApoJ�����ֶ�̬�ͣ���ApoJ1��ApoJ2��ApoJ3��ApoJ1�������������������ڡ�ApoJ2�������������������������˺��ˣ�����Ƶ�ʷֱ�Ϊ24%��28%��ApoJ3����һ�������������߲������ɴ˶�̬�Ե�ͻ����δ�������������֡���λ�����ƺ����ܵ��̴���LDL���̴���HDL���̴���VLDL���̴�������������ˮƽû��������Ӱ�졣
��������
�������ŷ�������ѧ����Ӧ����֬���л�붯������Ӳ�����о����������б���������֬����cDNA���������������й���֬�������̬�Եı������վ��������Ƕ�֬���״�л���Ҵӷ���ȱ�ݵ�ˮƽ������ʶȡ���˳���Ľ�չ��Ȼ����������֬��������������δ��ȫ����������������ApoE4�������Գմ��йأ���ʾ��ApoE���µĹ��þ���֤����һ�㡣�Ը�����֬���Ļ���ĸ��ֶ�̬�������������ķֲ�����ϵͳ�ļ����������д��ڽ�һ��Ŭ����һЩ�µġ����ġ�ȷ�ļ�ⷽ��Ҳ�д��ڿ�������Щ�о�����Ϊ��֬�����붯������Ӳ���������Ƶ�̽���Լ��Զ�������Ӳ������IJ��ķ���Ѱ���µĴ�ʩ�����µ�ǰ����
��Ҷ���壩
�����Ρ������ġ���
����1.Li WH, Tanzmura M, Luo CC, et al Datta, S,and Chan L, The apolipioprotein multigene��family: biosynthesis,structue,structure-function relationships, and evolution.J Lipid Res,1988,29:245-271
����2.Chan,L,The,apolipoprotein multigene family: structure,express evolution and molecular genetics.Kin.Wochenschr,1986,67:225-237
����3. Young S G.Recent progress in understanding apolipoprotein B.Circulation,1990,82:1574-1594
����4.Mclean JW ,Tomlinson JE,Kuang WJ,et al cDNA sequence of hunan apoliporpotein(a) is homologous to plasminogen.Nure 1987,330:132-137
����5. Ichinosie A.Multiple members of the plasminogen,apolipoprotein(a) gene family associated with thromboses.Biochemistry 1992,31:3113-3118
����6.Shoulders CC,Kornblihtt AR,Munro BS,et al Gene structure of human apolipoprotein AI.Nucleic Acid Res,1983,88;2827-2837
����7. Tsao YK,Wei CF,Robberson DL,et al.Isolation and chraacterization of the human apolipoprotein A��gene.Electron microsscoic analysis of RNA:DNA hybrids,nucleotide sequence ,identification of a polymorphic MspI site and general structural organization of apolipoprotein genes J Biol Chem,1985,260:15222-15231
����8. Karathanasis SK,Yunis I,Zannis,VI.Structure,evolution and tissue apecific synthesis of human apolipoprotein AIV.Bochem,1986,25:3962-3970.
����9.Blackhart BD,Ludwlg EM,Pierotti VR,et al Structure of the Human apolipoprtein B gene.j Biol Chem,1986,261:15264-3970
����10. Knott TJ,Robertson ME,Priestley LM,et.al Characterization of mRNAs encoding the precursor for human apoliporpotein CI.Nucleic Acid Res,1984,12:3909-3915
����11. Wei CF,Tsao YK,Robberson DL,et al.The structure of the human apolipopretinC�� gene.Electron microscopic analysis of RNA:DNA hybrids complete nucleotide sequence and identification of 5��homologous sequences among apolipoprotein genes J Biol Chem.1985,260:15211-15221
����12.a Protter AA,Levy Wilson B,Miller J,et al Isolation and sequence analysis of the human apolipoprotein C-��gene an dthe intergenic region between AopAI and ApoC-�� genes and the intergenic region between ApoAI and ApoC-�� genes..DNA,1984,3:449-456
����13.b Allan CM,Wallker D,Segrest JP,et al Identification and characterization of a new human gene (APOC-��)in the apolipoprotein E,C-I,and C-��gene locus .Genomics ,1995,28291-300
����14.Drayna DT,McLean JW,Wion KL,et al Human apolipoprotein Dgene:gene sequence,chromosome localization and homology to the ? globulin �Cglobulin superfamily ,DNA,1987,6:199-124
����15.Das HK,McPherson J,Bruns GAP,et al.Isolation and characterzation and mapping to chromosome 19 of the human apolipoprotein E.Cene,J Biol Chem,1985,260:6240-6247
����16. Day JR,Albers JJ,Gilbert TL,et al Purification and molecular cloning of human apolipoprotein F.Biochoim Biophgys Res Commun,1994,203:1146-1151
����17. Steinkasser A,Estaller C,Weiss EH,et al Complete nucleotide and deduced amion acid sequence of human human ��2-glycoprotein I.Biochem J,1991,227:387
����18.De Silva HV,Harmony JAK,Stuart WD,et al Apolipoprotein J:struture and tissue distrbution Biochem,1990,29:5380-5389
����19.Kao FT,Chan L.Ghromosomal fine mapping of apolipoprotein genes in the human genome by somatic cell hybrids Methods Enzymol,1986,128:851-863
����20.Harper ME,Chan L,Chromosomal fine mapping of apolipoprotein genes by in situmucleic acid hybridization to metaphase chromosomes Metthods Enzymol,1986,128:863-786
����21. Karathanasis SK .Apolipoprotein mutligene family and tandem organization of human apolipoprtein A-I,C-�� and A-�� genes .Proc Natl Acad Sci USA,1985,82:6374-6378
����22.Lusis AJ,Heinzmann C,Sparkes RS,et al.Regional mappoing of human chromosome 19:organization of genes for plasma lipid transport(APOC1,-C2,and-E and LDLR)and the genes C3,PEPD,and GPI.Proc Nat Acad Sci USA ,1986,83:3929-3933
����23. Spardes RS.Winokur S,Lusis AJ,et al Regional assignment of the apolipoprotein A I gene by in situ hybridization to human chromosome iiq-qter.Cytogenet Cell Genet,1987,46:697
����24 .Middleton-Price HR,Vanden Berghe JA,Scott J,et al.Regional chromosomal localisation of ApoA�� to lq21-lq23.Hum Genet,1988,79:283-285
����25. Law SW,Lee N,Monge JC et al Human ApoB100 gene resides in the P23��pter region of chromosome 2.Biochim Biophys Res Commun,1985,131:1003-1012
����26. Olaisen B,Teisberg P,Gedde-Dahl T,et al. The apolipoprotein E(ApoE)locus on human chromosome 19.Cytogenet Cell Genet,1984,37:559
����27. Steindasserer A,Cockbarn DJ,Black DM,et al.Assignment of apolipoprtoein H(ApoH:beta-2glycoprotein I)to human chromososme 17q23��qter;determination of the major expression site Cytogenet Cell Genet .1992,60:31-33
����28. Fink TM.Zimmer M.Tschopp J,et al Human chusterin(CLI)maps to 8p21 in proximity to the lipoprotein lipase(LPL)gene.Genomics,1993,16:526-528
����29. Walden CC,Hegele RA.Apolipoprotein E in Hyperlipdemia Am Intem Med,1994,120:1026-1036
����30.Kane JP,Havi RJ.Disorders of the biogenesis and secretion of lipoproteins containing the B apoliprproteins In:The metabolic and molecular bases of inherited disease eds Scriver,C,R,Bearder AL,Sly W S,and Valle D,7thedition ,Voll,1995,1583-1885,McGraw-Hill,Inc, New York,U.S.A
����31.Brom MS,Goldstein JL,A receptor-mediated pathway for cholesterol homeostasis ,Science,1986,232:34-47
����32. Soria LF,Ludwig EH,Clarke HRG,et al.Association between a specific apolipoprtein B mutation and familial defective apolipoprotein B-100,Proc Natl Acad Sci USA,1989,86:587-591
����33. Hegele RA,Huang LS,Herbert PN,et al Apolipoprotein B gene DNA polymorphipms associated with myocardial infarction .N Engl j Med,1986,315:1509-1515
����34.a.Visvik S,Cambou JP,Arveiler D et al.Apolipoprotein b signal peptide poly morphism in patients with my ocardial infartions and controls, Hum Genet,1993,90:561-565
����35.b.Kammerer CM,Vandeberg JL,Haffner SM,et al Apolipoprotein B(ApoB)signal peptide length polymorphisms are associated with ApoB.low density lipoprotein cholesterol and glucose levels in mexican Americans.Atherosclerosis,1996,120:37-45
����36.Kinjff PD,Van den Maagdenberg AMJM,Frants RR.et al Genetic heterogeneity of apolipoprotein Eand its in fluence on plasmea lipid and lipoprotein levels Hum Mutation,1994,4:178-194
����37. Mahley RW,Stanley Crall Jr,Type��hyperlipoproteinemia(Dysbetalipoproteinemia)The role of apolipoprotein Ein normal and abnormal ilpoprotein metabolism In:The metabolic and molecular bases of intherited disease,7thed eds Scriver C.R,Beaudet A L,Sly W S,and Valle D Vol ��,pp1953-1980,McGraw-Hill,Inc,New Your,1995
����38.Rose AD Apolipoprotein E alleles as risk factors in alzheimers disease .Am Rev Med ,1996,47:387-400
����39.Utermann G,Menzel HJ,Kraft HG,et al Lp(a) glycoprotein phenotypes ,Inheritance and relation to Lp(a)lipoprotein concentrations in plasma.Jclin Invest.1987.80:458-465
����40.Utermann G.Lipoprotein(a) In:The metabolic and molecular bases of inherited disease.7th ed ,Vol.��,eds .Scriver C R.Beandet A L.Sly W.S,Valle,D.pp1887-1912,McGraw-Hill,Inc,New Your,1995
����41.Scanu AM,Edelstein C.Kringle-dependent structural and functional polymorphism of apolipoprotein(a).Biochim Biophys Acta,1995,1256:1-12
����42. Mooser V,Mancini FP,Bopp S et al Sequence polymorphisms in the Apo(a)gene associated with specific levels of Lp(a)in plasma.Hum Mol Genet,1995,209:372-378
����43.Ichinose A,Kuriyama M.Detection of polymorphisms in the 5��flanding region of the Gene for apolipoprotein(a),Biochim biophys Res Commam,1995,209:372-378
����44. Breslow JL Apoipoprotein genes and atherosclerosis.Clin Investig,1992,70:377-384
����45.Von eckardstein A,Funke H.Walter M,et al.Structural analysis of human apolipoprotein AI variants Jbiol Chem,1990,265:8610-8617
����46.Talmud PJ,Ye S.Humphries SE,Polymorphism in the promotor region of the apolipoprotein AI gene associated with differences in apolipoprotein AI levels:The European Atherosclerosis Research Study.Genet Epidemiol,1994,11:265-80
����47.Saha N,Tay JS,Low PS,et al Guanidine to adenine(G/A)substitution in the promoter region of the apoliprotein AI gene is associated with elevated aerum apolipoprotein AI levels in Chinese non-smoders Genet Epikemiol,1994,11:255-64
����48.Wang XL.Badenhop R,Humphrey KE,et al.New Mspl polymorphism at +83 BP of the human apolipoprotein AI gene-Association with increased circulating high density lipoprotein cholesterol levels.Genet Epidemid,1996,13:1-10
����49.Humphries SE.DNA polymorphisms of the apolipoprotein genes-their use in the investigation of the genetic component of hyperlipidaemia and atherosclerosis.Atherosclerosis,1988,72:89-108
����50.Dallinga-Thie GM,Bu XD,Trip MV,LS,et al.Apolipoprotein A-��/C��/A-��gene cluster in familial combined hyperlipidemia:effects on LDL-cholesterol and LDL-cholesterol and apoplipoproteins B and C��,J Lipid Res,1996,37:136-147
����51.Klasen EC,Talmud PJ,Havekes L,et al.A common restriction fragment length polymorphism of the human apolipoprotein E gene and its relationship to type �� hyperlipidaemia Hum Genet,1987,75:244-247
����52.Deeb SS,Takata K,Peng RL,et al A splice-uunction mutation responsible for familial apolipoprotein A-�� deficiency Am J Hum genet,1990,46:822-827
����53.Zaiou M,Visvidis S,Gueguen R,et al DNA polymorphisms of human apolipoprotein A-�� gene frequency and effects on lipid,lipoprotein and apolipoprtein lkevels in a French population Clin Genet.1994,46:248-254
����54. Kamboh MI,Crawford MH,Aston CE Population distributions of ApoE,ApoH,and Apoa�� poly morphisms and their relationships with quantetative plasma lipid levels among the Evenki herders of Siberia .Hum Biol,1996,68:231-243
����55.Brunzell JD.Familial ilpoprotein lipase deficiency and other carses of the chylomicronemia syndrome .In:Themetabolic and molemlar bases of inherited disease.7th ed ,Vol ��,eds Scriver C R,Beandet A L,Sly W S,Valle D,pp1913-1932,McGraw-Hill,Inc,New Yourk,1995
����56.Beheshti I,Hanson NQ,Copeland KB,et al. Single strand conformational polymorphisms(SSCP):Studies of the genetic polymorphisms of exon4of apolipoprotein C��,Clin Biochem,1995,28:303-307
����57.li WW,Dammerman MM.Smith JD,et al Common genetic varation in the promotor of the human ApoC�� gene abolishes regulation by insulin and may contrbute to hypertriglyceridemia.J Clin Invest,1995,96:2601-2605
����58.Vijayaraghavan S,Hitman GA,Kopelman PG,Apolipoprotein-d polymorphism:A genetic marker for obesity and hyperinsulinemia.J Clin Endocrinol Metab,1994,79:568-570
����59.Roberts SB,Greenberg AS The new obesity genes .Nutrition Reviews ,1996,54:41-49
����60.Steindasserer A,Dorner C.Wurzner Ret al.Human ��-glcoprotein ��:molecular analysis of DNA and amino acid polymorphism.Hum Genet,1993,91:402-402
����61.Kamboh MI,Harmory JAK.Sepehrnia B,et al Genetic studies of human apolipoproteins XX.Genetic polymorphism of apolipoprtein J and its impact on quantitative lipid traits in normolipidemic subjects Am J Hum Genet,1991,49:1167-1173