二、影响实验动物免疫反应的因素
免疫反应是随着动物种的进化而复杂化、精密化和完善化的,不同品种和品系动物的免疫反应有很大差异,因此在免疫学研究中选择动物时要特别注意动物遗传因素的影响,还要注意动物的年龄因素、感染因素、营养因素、药物因素、母源因素、应激因素、免疫抑制剂等因素对动物免疫反应的影响。
(一)动物的遗传因素
根据研究表明小鼠、豚鼠、家兔等动物对特异性抗原的免疫反应受遗传控制。动物体内免疫反应的基因决定着动物对各种疾病的易感性,决定着自身免疫病和体液免疫反应。这种免疫反应的基因紧密连接在这些动物体内主要组织相容系统上。如带等位基因H-2b的小鼠(如C57BL,C57L,129/J)比带有等位基因H-2K的小鼠(如C58,AKR,C3H)的抵抗力强,后者对小鼠白血病病毒和肿瘤病毒十分易感,又如SWR/J(H-2q)小鼠对淋巴细胞性脉络丛脑膜炎病毒(LCM)非常敏感,而C3H/J(H-2k)小鼠对该病毒有强大的抵抗力。这些例子说明由于遗传因素的影响,不同品系动物的免疫反应是有明显差异的。此外,不同种类动物的免疫反映也有差异,如研究第Ⅳ型变态反应(Arthus反应),家兔是一种好的实验动物,而豚鼠和大鼠不能采用。豚鼠通常产生少量的IgM。
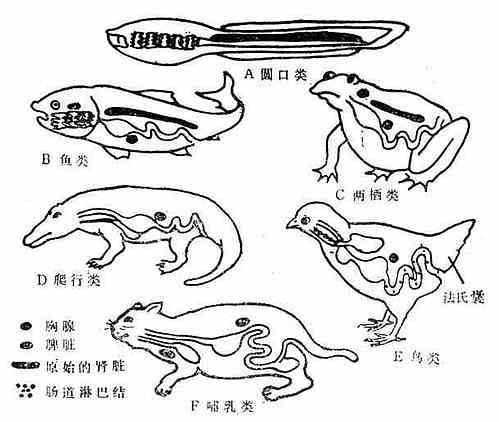
脊椎动物免疫系统的发生与种系化密切有关(见图10-1、2和表10-1):原始脊椎动物的淋巴器官发育还不完善,如圆口类动物沿其消化道有散在的淋巴结和淋巴细胞,并出现了胸腺,随着进化有了原始的肾脏,在鱼类还出现了肝脏。这些器官和组织开始时也多分布在消化道附近,这是由于原始脊椎动物(圆口类)及鱼类摄食时吸进大量水,并通过鳃孔将水排出,因此咽头部最先遭受病原微生物的侵击,为此在消化道附近产生了相应的御体系。而到了高等脊椎动物,由于种系的进化,这些器官的分布就多样化了。但从高等动物胸腺个体发生来看,它也是从第三、第四咽囊腹侧上皮演化发育而来,说明这跟种系发生有关。
虽然圆口类动物就能产生IgM,但无控制B淋巴细胞分化发育的中枢器官,至两栖类开始出现较原始的类囊器官。而爬行类动物的类囊,位于消化道的末端。鸟类以外其他动物有类体液免疫完善的免疫系统。
因此,在免疫学研究中进行实验动物选择时,要特别注意遗传因素对免疫反应的影响,各种实验动物具有不同的免疫反应和免疫特点。下面再举个典型例子。实验动物补体系统各成分的缺陷因实验动物的种类不同而有明显差异。
补体缺陷(Complemenr Deficiencies)
图10-1 脊椎动物的免疫反应
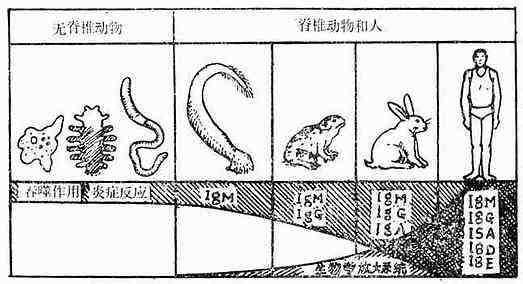
图10-2 免疫应答反应的种系发育主要代表
C1 鸡
C2 豚鼠、大鼠
C3 狗(Brittary Spaniel)
C4 金黄地鼠
C5 小鼠(K/HeN.AKR/N、B10、DZ/DsnN)
C6 兔、地鼠
当C1g缺乏时可出现严重的联合性免疫缺乏病,反复发生威胁生命的感染;C1r缺乏时发生坏疽性红斑,反复的细菌感染,狼疮样综合症;C1s缺乏时,出现红斑狼疮,进行性肾小球肾炎,关节炎;C4缺乏时可发生狼疮,关节炎、类过敏性紫癜;C2缺乏时发生狼疮,致死性皮肌炎,类过敏性紫癜,狼疮样综症,进行性肾小球肾炎,反复感染;C3缺乏时对感染的易感性升高;C5缺乏时可发生狼疮,腹泻及消耗病:C6缺乏时,可发生反复G(-)感染,淋菌性多关节炎,反复发生脑膜炎。
表10-1 脊椎动物免疫系统种系发生的比较
动 物 |
细 胞 |
淋 巴 器 官 |
免 疫 反 应 |
||||||||||
分类 |
动物 名称 |
淋巴细胞 |
浆细胞 |
胸腺 |
脾脏 |
淋巴结 |
法氏囊 |
细胞免疫 |
体 液 免 疫 |
||||
| IgM | IgG | IgA | IgE | IgD | |||||||||
圆口类 |
鳗 | + | - | + | - | - | - | + | + | - | - | - | - |
鱼类 |
油鲛 | + | - | + | + | - | - | + | + | - | - | - | - |
| 鲤鱼 | + | + | + | + | - | - | + | + | - | - | - | - | |
两栖类 |
蝾螈 | + | + | + | + | - | + | + | - | - | - | - | |
| 青蛙 | + | + | + | + | (+) | +* | + | + | + | - | - | ||
爬行类 |
龟蛇 | + | + | + | + | (+) | ? | + | + | + | - | - | - |
鸟类 |
鸡 | + | + | + | + | + | + | + | + | + | +? | - | - |
哺乳类 |
鼠、人 | + | + | + | + | + | +* | + | + | + | + | + | + |
(+)可能存在,+*有功能相似的类囊器官。

图10-3 各种动物胸导管淋巴细胞数目的变化(细胞数/公斤体重)
(二)动物的年龄因素
免疫学研究中选择适宜年龄的实验动物是非常重要的,年龄影响着实验动物的免疫机能。如幼龄动物的免疫系统机能发育不完善或机能很弱。一般来说,动物的免疫机能在青年期达到高峰,以后随着年龄增大逐渐减弱,主要表现有血清中免疫球蛋白含量低,细胞免疫功能下降,恶性肿瘤和自身免疫性疾病的发病率增高等。据研究,小鼠、大鼠和豚鼠随年龄增加免疫反应的活性也减弱,老龄鼠产生IgG和IgM的能力仅为年青成年鼠的10%左右,细胞免疫同样也减弱。因此,老龄鼠对诱发肿瘤极敏感。
实验证明给胚胎期或新生期的动物注射异基因型细胞时很容易造成对该细胞的免疫无反应性(Immunolgical Unresponsivenss),而对成年动物注射异基因型细胞时通常可引起免疫反应。据认为这种免疫耐受性与免疫系统(包括中枢淋巴样器官即骨髓、胸腺和外周淋巴样器官即脾脏、淋巴结、肠管相关淋巴样组织以及循环的淋巴样细胞)的发育未臻成熟有关。
Heidrick和Makindan氏认为老龄鼠细胞免疫中免疫细胞的缺乏,可能是由于T细胞和B细胞没有能力增殖的缘故。通常以为动物越是趋于老年,免疫反应的自隐机制被破坏的机会就越多,因而发自身免疫疫病的机会就越多。这种自稳对动物的正常生命活动乃至生存具有重要意义。在正常情况下,健康机体对自己的细胞、非细胞成分如蛋白质、多糖等是不发生“明显的”免疫反应的(自身免疫耐受状态,Selftolerance),而对非自己的抗原则发生免疫反应,也就是说机体通过免疫系统能识别“自己”和“非自己”。这种自我识别能力称之为自稳机制。
(三)动物的母源因素
各种哺乳类动物的胎儿和出生仔畜免疫的获得不同。在初乳中主要为IgA,初生动物血清中的母源抗体大部分为IgG,对仔畜预防病毒和细菌感染起着一定的保护性免疫作用。大部分IgA对粘膜表面起着局部保护作用,而IgG起着总的保护作用。但是母源抗体还有有害作用,它能诱发新生动物溶血病(如驹)和抑制出生动物的主动免疫。
雌性动物将母源抗体转移给胎儿或仔畜的途径和特异性不同,这与动物胎盘的结构和类型有关(见下页表10-2),一般来说,有三种主要的转移途径,随动物种类不同,有的经绒毛膜尿囊胎盘转移;有的经卵黄囊上皮和卵黄循环转移;还有是初乳经肠道吸收。前两者被认为是胎儿期获得抗体的途径,后者为出生后转移抗体的途径。
免疫球蛋白的转送是有选择性的,有些种类的抗体易转移,同种(系)抗体转移比异种抗体快。胎盘对各种母源抗体也有选择性,如:灵长目中Igg 易通过胎盘屏障,IgM、IgA和IgE则不能。家兔的IgG和IgM易通过胎盘到达胎儿。关于母源抗体选择性的转移机制还不清楚,一般认为,这种选择性的转移是由于组织能选择性地识别抗体的F。端造成的,母源抗体转移中具有选择作用的组织见表10-3。动物常乳和初乳中检出抗体的类型见表10-4。
表10-2 动物的胎盘屏障和母源抗体的转移
| 动 物 | 胎盘类型 |
结构 (层数) |
转移途径 | 选择性 |
抗体转移 |
持续时间 |
|
出生前 |
出生后 |
||||||
小鼠 |
血性绒毛膜 | 4 |
肠(主要)卵黄囊 | + |
+(卵黄囊) | 艹 |
出生到17天 |
大鼠 |
血性绒毛膜 | 4 |
肠(主要)、卵黄囊 | + |
+(卵黄囊) | 廿 |
出生到20天 |
豚鼠 |
血性绒毛膜 | 2 |
卵黄囊(主要)、肠 | ? |
卅 | + |
整个妊娠期间和出生后2天内 |
家兔 |
血性绒毛膜 | 2 |
卵黄囊 | + |
卅 |
- |
从妊娠开始15天起 |
灵长类 |
血性绒毛膜 | 2 |
胎盘 |
+ |
卅 |
卅 |
妊娠开始3个月 |
猬 |
肠 |
+ |
- |
艹 |
出生到35天 | ||
狗 |
内皮绒毛膜 | 4 |
肠 |
? |
- |
卅 |
出生到8天 |
猫 |
肠 |
? |
- |
卅 |
不定 | ||
水貂 |
肠 |
? |
- |
卅 |
出生到8天 | ||
雪貂 |
肠 |
? |
- |
卅 |
出生到34天 | ||
猪 |
上皮绒毛膜 | 6 |
肠 |
- |
- |
卅 |
出生后36小时 |
马 |
上皮绒毛膜 | 6 |
肠 |
- |
- |
卅 |
出生后36小时 |
牛 |
结缔组织绒毛膜或上皮绒毛膜 | 6 | 肠 | - |
- |
卅 |
出生后36小时 |
绵羊 |
结缔组织绒毛膜或上皮绒毛膜 | 6 |
肠 |
- |
- |
卅 |
出生后36小时 |
表10-3 母源抗体转移中呈现选择作用的组织
动 物 种 类 |
组 织 |
灵 长 目 |
胎 盘 |
大鼠与小鼠 |
肠 |
家 兔 |
卵 黄 囊 |
偶 蹄 兽 |
乳 腺 |
鸟 |
卵黄囊膜 |
表10-4 动物常乳和初期中免疫球蛋白类型
动 物 |
常乳和初乳中免疫球蛋白类型 |
|
家 兔 |
初乳 |
IgA |
灵长目 |
初乳 |
IgA |
大 鼠 |
初乳 常乳 |
IgA(多量)IgGa,IgGb,r1,IgM(很少) |
| IgG(多量) | ||
小 鼠 |
初乳 |
IgA,快和慢的IgG |
狗 |
初乳 常乳 |
IgG(多量),IgA,IgM |
| IgA(多量),IgG,IgM | ||
猪 |
初乳 常乳 |
IgG(多量),IgA,IgM |
| IgA(多量) | ||
牛 |
初乳 |
快IgG(r1)IgM(β2) |
绵 羊 |
初乳 |
快IgG(r1),IgA |
(四)动物的感染因素
病毒、细菌、真菌或寄生虫的感染都能多方面地改变动物机体的生理功能,这种改变无疑将影响到动物的免疫系统。动物感染可引起继发性营养不良,从而影响到免疫反应。一般实验动物常发生病毒的隐性感染,在以下几方面影响免疫功能:抗体产生的方式,免疫球蛋白的数量,免疫耐受性的产生,植皮排斥,植皮对宿主的反映,迟发型变态反应,淋巴细胞转化和吞噬作用等。
动物病毒感染引起的免疫抑制表现:干扰正常免疫系统的功能;改变抗原的吸收和处理;破坏抗体形成细胞和浆细胞(如白血病病毒)。病毒感染后也可引起免疫增强的表现,如乳酸脱氨酶病毒LDH,委内瑞拉马脑脊髓炎病毒VEE等病毒,能增加产生抗体细胞数量:感染病毒的细胞能产生微量核苷酸,起着佐剂的作用;病毒还能促进免疫细胞对抗原的处理。
动物感染呼吸道病毒、腺病毒、流行性脊髓灰质炎病毒、仙台病毒、痘苗病毒、单纯疮疹病毒、新城疫病毒等病毒时,能够抑制淋巴细胞的转化。肝炎、肢骨发育不全(Ectromelia)、淋巴细胞性脉络丛脑膜炎(LCM)、乳酸脱氨酶病毒(LDH)能抑制网内皮系统的机能。而Friend白血病、Moloney白血病和委内瑞拉马脑脊髓炎(VEE)能增强吞噬细胞的吞噬作用。