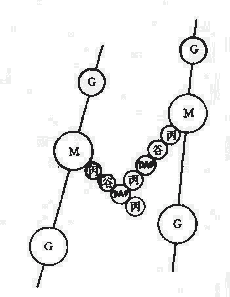
第二节 细菌的结构
细菌的结构对细菌的生存、致病性和免疫性等均有一定作用。细菌的结构按分布部位大致可分为:表层结构,包括细胞壁、细胞膜、荚膜;内部结构包括细胞浆、核蛋白体、核质、质粒及芽胞等;外部附件,包括鞭毛和菌毛。习惯上又把一个细菌生存不可缺少的,或一般细菌通常具有的结构称为基本结构,而把某些细菌在一定条件下所形成的特有结构称为特殊结构(图2-2)。
图2-2 细菌的结构
一、基本结构
细菌基本结构包括细胞壁、细胞膜、细胞浆及核质。
(一)细胞壁(Cell wall)细胞壁为细菌表面比较复杂的结构。是一层较厚(5~80nm)、质量均匀的网状结构,可承受细胞内强大的渗透压而不破坏。细胞壁坚韧而有弹性。
1.细胞壁主要组份:主要成分是肽聚糖(Peptidoglycan),又称粘肽(Mucopetide)。细胞壁的机械强度有赖于肽聚糖的存在。合成肽聚糖是原核生物特有的能力。肽聚糖是由N-乙酰葡萄糖胺和N-乙酰胞酸两种氨基糖经β-1.4糖苷键连接间隔排列形成的多糖支架。在N-乙酰胞壁酸分子上连接四肽侧链,肽链之间再由肽桥或肽链联系起来,组成一个机械性很强的网状结构(图2-3)。各种细菌细胞壁的肽聚糖支架均相同,在四肽侧链的组成及其连接方式随菌种而异。
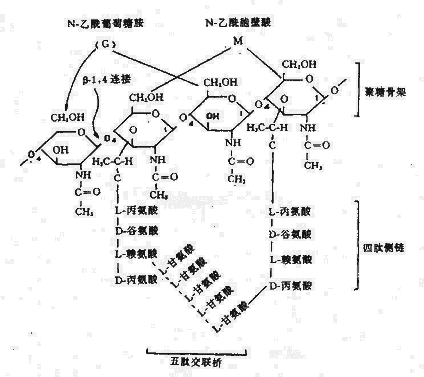
图2-3 肽聚糖分子结构
革兰氏阳性菌例如葡萄球的四肽侧链氨基酸由D-丙-D-谷-r-L-赖-D-丙组成。初合成的肽链末端多一个D-丙氨酸残基。肽桥是一条5个甘氨酸的肽链,交联时一端与侧链第三位上赖氨酸连接,另一端在转肽酶的作用下,使另一条五肽侧链末端D-丙氨酸脱去,而与侧链第四位D-丙氨酸连接。从X光检查可见肽聚糖的多糖链是一条较硬而又呈螺旋状卷曲的长杆,由于其呈螺旋状,连接在其上的肽链才伸向四方,使交联受到一定了限制,只有邻近的肽链才可交联。但葡萄球菌的肽桥较长,有可塑性,使远距离的肽链间也可交联,交联率达90%,形成坚固致密的三维立本网状结构(图2-4)。
而革兰氏阴性大肠杆菌的四肽侧链中第三位的氨基酸被二氨基庚二酸(DAP)所取代,以肽链直接与相邻四肽侧链中的D-丙氨酸相连,且交联率低,没有五肽交联桥,形成二维平面结构,所以其结构较革兰氏阳性的葡萄球疏桦(图2-5)。
|
|
图2-4 金黄色葡萄球糖细胞 壁的肽聚糖结构 |
图2-5 大肠杆菌细胞壁的肽聚糖结构 |
| M:N-乙酰胞酸 | G:N-乙酰氨基葡萄糖 | O:β-1,4糖苷链 | a:L-丙氨酸 |
| b:D-谷氨酸 | c:L-赖氨酸 | d:D-丙氨酸 | x:甘氨酸 |
凡能破坏肽聚糖结构或抑制其合成的物质,都能损伤细胞壁而使细菌变形或杀伤细菌,例如溶菌酶(Lysozyme)能切断肽聚糖中N-乙酰葡萄糖胺和N-乙酰胞壁酸之间的β-1.4糖苷键之间的联苷键之间的联结,破坏肽聚糖支架,引起细菌裂解。青霉素和头孢菌素能与细菌竞争合成胞壁过程所需的转肽酶,抑制四肽侧链上D-丙氨酸与五肽桥之间的联结,使细菌不能合成完整的细胞壁,可导致细菌死亡。人和动物细胞无细胞壁结构,亦无肽聚糖,故溶菌酶和青霉素对人体细胞均无毒性作用。除肽聚糖这一基本成份以外,革兰氏阳性菌和革兰氏阴性菌还各有其特殊结构的成分。
2.革兰氏阳性菌细胞壁特殊组份细胞壁较厚,约20~80mm。肽聚糖含量丰富,有15~50层,每层厚度1nm,约占细胞壁干重的50~80%。此外,尚有大量特殊组份磷壁酸(Teichoic acid)。 (图2-6)磷壁酸是由核糖醇(Ribitol)或甘油(Glyocerol)残基经由磷酸二键互相连接而成的多聚物。磷壁酸分壁磷壁酸(Wall teichoic acid)和膜磷壁酸(Membrane teichoic acid)两种,前者和细胞壁中肽聚糖的N-乙酰胞壁酸连结,膜磷壁酸又称脂磷壁酸(Lipteichoic acid)和细胞膜连结,另一端均游离于细胞壁外。磷壁酸抗原性很强,是革兰氏阳性菌的重要表面抗原;在调节离子通过粘肽层中起作用;也可能与某些酶的活性有关;某些细菌的磷壁酸,能粘附在人类细胞表面,其作用类似菌毛,可能与致病性有关。
此外,某些革兰氏阳性菌细胞壁表面还有一些特殊的表面蛋白,如A蛋白等,都与致病有关。
3.革兰氏阴性菌细胞壁特殊组份细胞壁较薄,约10~15nm,有1~2层肽聚糖外,约占细胞壁干重的5~20%。结构比较复杂。尚有特殊组份外膜层位于细胞壁肽聚糖层的外侧,包括脂多糖、脂质双层、脂蛋白三部分(图2-6)。
脂蛋白(Lipoprotein)一端以蛋白质部分共价键连接于肽聚糖的四肽侧链上另一端以脂质部分经共价键连接于外膜的磷酸上。其功能是稳定外膜并将之固定于肽聚糖层。

图2-6 细菌细胞壁结构模式图脂质双层是革兰阴性菌细胞壁的主要结构,除了转运营养物质外,还有屏障作用,能阻止多种物质透过,抵抗许多化学药物的作用,所以革兰氏阴性菌对溶菌酶、青霉素等比革兰氏阳性具有较大的抵抗力。一些化学物质如乙二胺四乙酸(EDTA)与2%十二烷基硫酸钠(SDS)或45%酚水溶液可以将外膜除去,而留下坚韧的肽聚糖层。此外,外膜蛋白质还可作为某些噬菌体和性菌毛的受体。
脂多糖(Lipopolysacchride,LPS)由脂质双层向细胞外伸出,包括类脂A、核心多糖、特异性多糖三个组成部分,习惯上将脂多糖称为细菌内毒素。
①类脂A:为一种糖磷脂,是由焦磷酸键联结的氨基葡萄糖聚二糖链,其上结合有各种长链脂肪酸。它是脂多糖的毒性部分及主要成份。为革兰氏阴性菌的致病物质。无种属特异性,各种革兰氏阴性菌内毒性引起的毒性作用都大致相同。
②核心多糖:位于类脂A的外层,由已糖、瘐糖、2-酮基―3―脱氧辛酸(KDO)、磷酸乙醇胺等组成。经KDO与类质A共价联结。核心多糖具有属特异性,同一属细菌的核心多糖相同。
③特异性多糖:在脂多糖的最外层,是由数个至数十个低聚糖(3~5单糖)重复单位所构成的多糖链。革兰氏阴性菌的菌体抗原(O抗原)就是特异多糖。各种不同的革兰氏阴性菌的特异性多糖种类及排列顺序各不相同,从而决定了细菌抗的特异性。
革兰氏阳性菌和革兰氏阴性菌的细胞壁结构显著不同,导致这两类细菌在染色性、抗原性、毒性、对某些药物的敏感性等方面的很大差异。
表2-1 革兰氏阳性菌与革兰氏阴性菌细胞壁结构的比较
特 征 |
革兰氏阳性菌 |
革兰氏阴性菌 |
强度 |
较坚韧 |
较疏松 |
厚度 |
厚,20~80nm |
薄,5~10nm |
肽聚糖层数 |
多,可达50层 |
少,1~3层 |
肽聚糖含量 |
多,可占胞壁干重50~80% |
少,占胞壁干重10~20% |
磷壁酸 |
+ |
- |
外膜 |
- |
+ |
结构 |
三维空间(立体结构) |
二维空间(平面结构) |
4.细胞壁的功能细菌细胞壁坚韧而富有弹性,保护细菌抵抗低渗环境,承受世界杯内的5~25个大气的渗透压,并使细菌在低渗的环境下细胞不易破裂;细胞壁对维持细菌的固有形态起重要作用;可充许水分及直径小于1nm的可溶性小分子自由通过,与物质交换有关;细胞壁上带有多种抗原决定簇,决定了细菌菌体的抗原性。
5.L型细菌 L型是指细菌发生细胞壁缺陷的变型。因其首次在Lister研究所发现。故以其第一个字母命名。当细菌细胞壁中的肽聚糖结构受到理化或生物因素的直接破坏或合成被抑制这种细胞壁受损的细菌一般在普通环境中不能耐受菌体内部的高渗透压而将胀裂死亡;但在高渗环境下,它们仍可存活而成为细菌细胞壁缺陷型。革兰氏阳性菌L型称为原生质体(protoplast),必须生存于高渗环境中。革兰氏阴性菌L型称为原生质球(spheroplast),在低渗环境中仍有一定的抵抗力。
细菌L型的形态因缺失细胞壁而呈高度多形性,有球状、杆状和丝状。大小不一,L型细菌大多数染成革兰氏阴性。细菌L型生长繁殖时的营养要求基本与原菌相同,但必须补充3%~5%的NaC1、10%~20%蔗糖或7%聚乙烯吡咯酮(PVP)等稳定剂,以提高培养基的渗透压。同时还需要加入人或马血清。L型细菌生长较缓慢,一般培养2~7天后在软琼脂平板形成中同较厚、四周较薄的荷包蛋样细小菌落。此外,L型菌尚有颗粒型和丝状型两种类型。L型细菌在液体培养基中生长后呈较疏松的絮状颗粒,沉于管底,培养液则澄清。
人工诱导或自然情况下,细菌L型在体内或体外均能产生。各种细菌L型有一个共同的致病特点。即引起多组织的间质性炎症。细菌变为L型致病性有所减弱,但在一定条件下L型又可复为细菌型,引起病情加重。变形后的细菌其形态、培养特性均发生了改变,以致查不出病原使许多病人贻误诊治。临床遇有症状明显而标本常规细菌培养阴性者,应考虑细菌L型感染的可能性,宜作细菌L型的专门培养。
(二)细胞膜(Cell membrane)或称胞膜(Cytoplasmic membrane)位于细胞壁内侧,包绕在细菌胞浆外的具有弹性的半渗透性脂质双层生物膜。主要由磷脂及蛋白质构成,膜不含胆固醇是与真核细胞膜的区别点。细胞膜有选择性通透作用,与细胞壁共同完成菌体内外的物质交换。膜上有多种呼吸酶,参与细胞的呼吸过程。膜上有多种合成酶,参与生物合成过程。细菌细胞膜可以形成特有的结构。
1.中介体(Mesosome)用电子显微镜观察,可以看到细胞膜向胞浆凹陷折叠成囊状物,称为中介体。中介体与细胞的分裂、呼吸、胞壁合成和芽胞形成有关。中介体位置常在菌体的侧面或靠近中央横隔处。横隔中介体与核质相连,当细菌分裂时横隔中介体也一分为二,各自带一套核质进入子代细胞;中介体扩大了细胞膜的表面积,相应地增加呼吸酶的含量,可为细菌提供大量能量,有拟线粒体(Chondroid)之称,中介体多见于革兰氏阳性菌。
2.胞质间间隙在革兰氏阴性细菌的细胞膜与细胞壁之间有一空间,称为胞质间间隙(Periplasmic space)。此处聚集了若干种胞外酶,主要是水解酶,与营养物质的分解、吸收和运转有关。能破坏某些抗生素的酶(如青霉素酶)亦集中在此间隙内。
(三)胞浆(Cytoplasm)是无色透明胶状物,基本成份是水、蛋白质、脂类、核酸及少量无机盐。细胞浆中还存在一些胞浆颗粒。
1.质粒(Plasmid)这是染色体外的遗传物质,为双股环状DNA。分子量比染色体小,可携带某些遗传信息,例如耐药因子、细菌素及性菌毛的基本均编码在质粒上。质粒能进行独立复制,失去质粒的细菌仍能正常存活。质粒可通过接合、转导作用等将有关性状传递给另一细菌。
2.核糖体(Ribosome)电镜下可见到胞浆中有大量沉降系数为70S的颗粒,即核糖体。其化学组成70%为RNA,30%为蛋白质。细胞中约90%的RNA和40%的蛋白质存在于核糖体中。当mRNA连成多聚核蛋白体(Polyribosome),就成为合成蛋白质的场所。细菌的70S核糖体由50S和30S两个亚基组成。链霉素能与细菌核糖体的30S基结合,红霉素能与50S亚基结合,从而干扰细菌蛋白质的合成而导致细菌的死亡;真核细胞的核糖体为80S,因此对人体细胞则无影响。
3.胞浆颗粒(Cytoplasma granula)大多数为营养贮藏物,较为常见的是贮藏高能磷酸盐的异染颗粒(Metachrometic granula),嗜碱性较强,用特殊染色法可以看得更清晰。根据异染颗粒的形态及位置,可以鉴别细菌。
(四)核质(Nnclear materal)或拟核(Nucleoid)是细菌的遗传物质,决定细菌的遗传特征。集中在细胞浆的某一区域,多在菌体中部。它与真核细胞的细胞核不同点在于四周无核膜,故不成形,也无组蛋白包绕。一个菌体内一般含有1~2个核质。现已证明,细菌的核质是由双股DNA组成的单一的一根环状染色体反复回旋盘绕而成,细菌的染色体是裸露的DNA。
大肠杆菌的染色体分子量为3×109,伸展后长度约达1.1mm,约含5×106碱基对,足可携带3,000~5,000个基因,以满足细菌生命活动的全部需要,核质具有细胞核的功能,控制细菌的各种遗传性状。细菌胞浆中含有大量RNA,用碱性染料染色着色很深,将核质掩盖,不易显露。若用酸或RNA酶处理,使RNA水解,再用富尔根(Feulgen)氏法染色,便可染出核质,在普通光学显微镜下可以看见,一般呈球状、棒状或哑铃状。
二、特殊结构
细菌的特殊结构包括荚膜、鞭毛、菌毛和芽胞。
(一)荚膜(Capsule)许多细菌胞壁外围绕一层较厚的粘性、胶冻样物质,其厚度在0.2um以上,普通显微镜可见,与四周有明显界限,称为荚膜。如肺炎双球菌(图2-7)。其厚度在0.2um以下者,在光学显微镜下才不能直接看到,必须以电镜或免疫学方法才能证明,称为微荚膜(Microcapsule),如溶血性链球菌的M蛋白、伤寒杆菌的Vi抗原及大肠杆菌的K抗原等。

图2-7 肺炎球菌荚膜
大多数细菌(如肺炎球菌、脑膜炎球菌等)的荚膜由多糖组成。链球菌荚膜为透明质酸;少数细菌的荚膜为多肽(如炭疽杆菌荚膜为D-谷氨酸的多肽)。
细菌一般在机体内和营养丰富的培养基中才能形成荚膜。有荚膜的细菌在固体培养基上形成光滑型(S型)或粘液型(M)菌落,失去荚膜后菌落变为粗糙型(R)。荚膜并非细菌生存所必需,如荚膜丢失,细菌仍可存活。
荚膜除对鉴别细菌有帮助外,还能保护细菌免遭吞噬细胞的吞噬和消化作用,因而与细菌的毒力有关。荚膜抗吞噬的机理还不十分清楚,可能由于荚膜粘液层比较光滑,不易被吞噬细胞捕捉之故。荚膜能贮留水分使细菌能抗干燥,并对其他因子(如溶菌酶、补体、抗体、抗菌药物等)的侵害有一定抵抗力。
(二)鞭毛(Flagllum)在某些细菌菌体上具有细长而弯曲的丝状物,称为鞭毛。鞭毛的长度常超过菌体若干倍。不同细菌的鞭毛数目、位置和排列不同,可分为单毛菌(Monotrichate)、双毛菌(Amphitrichate)、丝毛菌(Lophotrichate)、周毛菌(Peritrichate)(图2-8)。

图2-8 细菌的鞭毛(示意)
鞭毛自细胞膜长出,游离于细胞外。用电子显微镜研究鞭毛的超微结构,发现鞭毛的结构分为:基础小体、钩状体和丝状体三个部分组成(图2-9)。
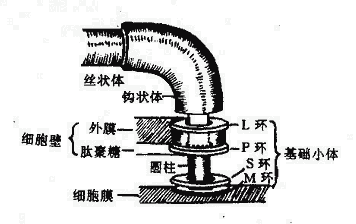
图2-9 大肠杆菌鞭毛根部结构模式图
1.基础小体(Basalbody)位于鞭毛根部,埋在细胞壁中。革兰氏阴性菌鞭毛的基础小体由一根圆柱和两对同心环所组成,一对是M环与S环,附着在细胞膜上;另一对是P环与L环,连在胞壁的肽聚糖和外膜上(M、S、P、L分别代表细胞膜、膜上、肽聚糖、外膜中的脂多糖)。革兰氏阳性菌的细胞壁无外膜,其鞭毛只有M与S环而无P环和L环。鞭毛运动需要能量,细胞膜中的呼吸链可供其所需。
2.钩状体(Hook)位于鞭毛伸出菌体之处,呈钩状弯曲,鞭毛此转变向外伸出,成为丝状体。
3.丝状体(Filament)呈纤丝状,伸出于菌体之外,是由鞭毛蛋白亚单位呈紧螺旋状缠绕而成的中空的管状结构。鞭毛蛋白是一种纤维蛋白,其氨基酸组成与骨骼肌动蛋白相似,可能与鞭毛的运动性有关。
鞭毛是细菌的运动器官,往往有化学趋向性,常朝向有高浓度营养物质的方向移动,而避开对其在害的环境。常存在于杆菌及弧菌中。鞭毛的数量、分布可用以鉴别细菌。鞭毛抗原有很强的抗原性,通常称为H抗原,对某些细菌的鉴定、分型及分类具有重要意义。
(三)菌毛(Pilus)菌毛是许多革兰氏阴性菌菌体表面遍布的比鞭毛更为细、短、直、硬、多的丝状蛋白附属器,也叫做纤毛(Fimbriae)。其化学组成是菌毛蛋白(Pilin),菌毛与运动无关,在光镜下看不见,使用电镜才能观察到。菌毛可分为普通菌毛(Commonpilus)和性菌毛(Sexpilus)两种。
1.普通菌毛 长0.3~1.0um,直径7nm。具有粘着细胞(红细胞、上皮细胞)和定居各种细胞表面的能力,它与某些细菌的的致病性有关。无菌毛的细菌则易被粘膜细胞的纤毛运动、肠蠕动或尿液冲洗而被排除,失去菌毛,致病力亦随之丧失。
2.性菌毛 有的细菌还有1~4根较长的性菌毛,比普通菌毛而粗,中空呈管状。性菌毛由质粒携带的一种致育因子(Ferility factor)的基因编码,故性菌毛又称F菌毛。带有性菌毛的细菌称为F+菌或雄性菌,无菌毛的细菌称为F-菌或雌性菌。性菌毛能在细菌之间传递DNA,细菌的毒性及耐药性即可通过这种方式传递,这是某些肠道杆菌容易产生耐药性的原因之一。
(四)芽胞(Spore)在一定条件下,芽胞杆菌属(如炭疽杆菌)及梭状芽胞杆菌属(如破伤风杆菌、气性坏疽病原菌)能在菌体内形成一个折光性很强的不易着色小体,称为内芽胞(Endospore),简称芽胞。
芽胞一般只在动物体外才能形成,并受环境影响,当营养缺乏,特别是碳源、氮源或磷酸盐缺乏时,容易形成芽胞。不同细菌开成芽胞还需不同的条件,如炭疽杆菌须在有氧条件下才能形成芽胞。成熟的芽胞可被许多正常代谢物如丙氨酸、腺苷、葡萄糖、乳酸等激活而发芽,先是芽胞酶活化,皮质层及外壳迅速解聚,水分进入,在合适的营养和温度条件下,芽胞的核心向外生长成繁殖体,开始发育和分裂繁殖。芽胞并非细菌的繁殖体,而是处于代谢相对静止的休眠休态,以维持细菌生存的持久体。
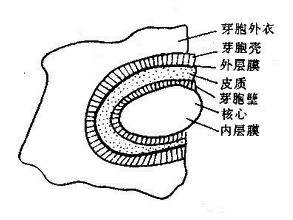
芽胞含水量少(约40%),蛋白质受热不易变性。芽胞具有多层厚而致密的胞膜,由内向外依次为核心、内膜、芽胞壁、皮质、外膜、芽胞壳和芽胞外衣(图2-10)。特别是芽胞壳,无通透性,有保护作用,能阻止化学品渗入。芽胞形成时能合成一些特殊的酶,这些酶较之繁殖体中的酶具有更强的耐热性。芽胞核心和皮质层中含有大量吡啶二羧酸(Dipicolinic acid,DPA),占芽胞干重的5~15%,是芽胞所特有的成分,在细菌繁殖体和其他生物细胞中都没有。DPA能以一种现尚不明的方式,使芽胞的酶类具有很高的稳定性。芽胞形成过程中很快合成DPA,同时也获得耐热性。
|
|
图2-10 芽胞结构模式 |
图2-11 细菌的芽胞形态 |
芽胞呈圆形或椭圆形,其直径和在菌体内的位置随菌种而不同,例如,炭疽杆菌的芽胞为卵圆形、比菌体小,位于菌体中央;破伤风杆菌芽胞正圆形、比菌体大,位于顶端,如鼓槌状。这种形态特点有助于细菌鉴别(图2-11)。芽胞在自然界分布广泛,因此要严防芽胞污染伤口、用具、敷料、手术器械等。芽胞的抵抗力强,对热力、干燥、辐射、化学消毒剂等理化因素均有强大的抵抗力,用一般的方法不易将其杀死。有的芽胞可耐100℃沸水煮沸数小时。杀灭芽胞最可靠的方法是高压蒸汽灭菌。当进行消毒灭菌时往往以芽胞是否被杀死作为判断灭菌效果的指标。
刘利兵