第二节 蛋白酪氨酸激酶
蛋白酷氨酸激酶(protein tyrosine kinase,PTK)是一类催化ATP上γ-磷酸转移到蛋白酪氨酸残基上的激酶,能催化多种底物蛋白质酪氨酸残基磷酸化,在细胞生长、增殖、分化中具有重要作用。迄今发现的蛋白酪氨酸激酶中多数是属于致癌RNA病毒的癌基因产物,也可由脊椎动物的原癌基因产。根据PTK是否存在于细胞膜受体可将其分成非受体型和膜受体型。
1.非受体型 以src基因产物为代表,此外还有Yes、Fyn、Lck、Fgr、Lyn、Fps/Fes及Ab1等。徐后两者外,其余非受体型蛋白酪氨酸激酶src家族分子理约为60kDa的蛋白质,它们之间除了N末端80个氨基酸组成不同外,其作部分都非常相似。
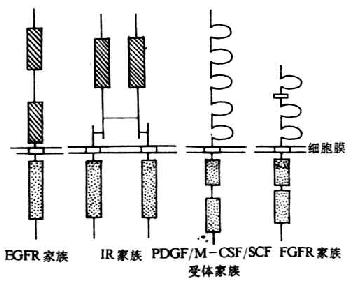
2.受体型 根据它们的结构不同,受体型酪氨酸激酶可以分为9种类型,其中较常见的有4种类型(图8-5)。
(1)表皮生长因子受体(EGFR)家族:EGF-R家族成员包括EGF-R(分子量为170kDa,广泛表达于多种组织细胞中)、erbB2/neu 及erbB-3基因表达产物。其家族成员的特点是在胞膜外有两个富含半胱氨酸的区域,胞浆内含有一个有酪氨酸激酶活化性的区域。
(2)胰岛素受体家族:其家族成员包括胰岛素受体(insulin receptor,IR)、胰岛素样生长因子-1受体(insulin-like growth factor-1receptor,IGF-1R)以及胰岛素相关受体(insulin related receptor,IRR)。胰岛素受体家族成员是由二个α亚单位和二个β亚单位通过链间二硫键形成的异源四聚体。其中α亚单位为配体结合部位;β亚单位的胞浆内部分含有酪氨酸激酶活性区域。
(3)PDGF/MCSF/SCF受体家族:其家族成员包括血小板衍生的生长因子α受体(PDGF-αR)、PDGF-βR、巨噬细胞集落刺激因子受体(M-CSFR)以及干细胞生长因子受体(SCFR)。以上成员的特点是胞膜外含有5个免疫球蛋白样结构域,胞浆内含有两个呈串联结构的酪酸激酶功能区。
(4)成纤维细胞生长因子受体(FGFR)家族:FGFR家族成员有FGFR1、FGFR2、FGFR3以及FGF4。它们的特点是在胞膜外含有3个免疫球蛋样结构域,其中在第1和第2结构域之间有一个含8个连续的酸性氨基酸结构,又称酸性盒结构域(acid box domain);胞浆内含有两个呈串联结构的酷氨酸激酶功能区。
图8-5 受体型酪氨酸激酶示意图
家体型酪氨酸蛋白激酶所介导的信号传递途径中涉及到一种重要的蛋白激酶即丝裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)。MAPK属于一种Ser/Thr蛋白激酶,可在多种不同的信号转导途径中充当一种共同的信号转导成份,且在细胞周期调控中发挥重要的作用。目前MAPK家族中至少有4个成员已被纯化和深入研究。如p42mapk,p44erk1,p54MAPK及p44mpk。这些MAPK最初由于不同的研究目的而被发现,因而它们各自还有着其它一些名称。如p42mapk也称为微管相关蛋白激酶(microtubule-associated protein-2 kinase,MAP-2k)及细胞外信号调节的激酶2(extracellular signal-regulated kinase2,ERK2)等。
属于src家族的蛋白酪氨酸激酶是一组膜结合蛋白,包括p60arc、p56lck、p59fyn、p59yes、(p62ues)、p56lyn、p59hck、p55fgr和p55blk等成员。但由于src家族PTKs缺乏胞外及跨膜序列,因此这些src家族PTKs通过其N末端与细胞膜表面蛋白的胞浆内结构域相连接,从而发挥信号转导作用(表8-2)。已证实p56lck参加CD4和CD8介导的信号传递;p59fyn参加TCR/CD3复合体介导的信号传递;p56lyn参加B细胞受体(BCR)介导的信号传递。经TCR途径T细胞活化后最早生化事件是几种内源性底物酪氨酸磷酸化水平的升高。这种酪氨酸磷酸化可能由p56kk或一个与PTK相关的称为p59fyn所介导。p59fyn主要表达于T淋巴中,发挥某些独特的调节功能。然而,还有一些调节功能更为广泛的PTKs表达于包括T细胞在内的多种细胞类型中,包括:(1)最初从小鼠脾细胞中发现的tk1蛋白产物;(2)在T细胞有丝分裂中数量可见增加并同T细胞瘤的发生有关的p34pim;(3)表达于T细胞中c-abl、ltk、c-kit和其它原癌基因蛋白产物。这些PTKs的确切功能还不清楚。
表8-2 同src家族PTKs相连接的细胞表面蛋白
| 细胞表面蛋白 | 细胞类型 | src家族PTK |
| TCR-CD3 | T | Fyn |
| CD2 | T | Fyn |
| CD4 | T | Lck |
| CD8 | T | Lck |
| CD45 | T | Fyn、Lck |
| BCR | B | Blk、Lyn(Fyn)、Lck |
| FcγRⅢ(CD16) | NK | Fyn |
| IL-2R | T,NK | Lck、Lyn |
| FcεR I | 肥大细胞、嗜碱性粒细胞 | Yes、Lyn(src) |
| FcεR Ⅱ(CD23) | 肥大细胞、嗜碱性粒细胞 | Fyn |
| CD36 | 血小板 | Lyn、Fyn、Yes |
一、p56lck
p56lck属于以p60v-src/c-src为代表的蛋白酪氨酸激酶家族成员。同这个家族的其它成员一样都具有酪氨酸激酶活性。目前发现,p56lck相对特异地存在于淋巴细胞中,尤其是成熟的静止T淋巴细胞中。p56lck可能在T细胞活化信号转导以及分化调节过程中起着重要的作用。
(一)p56lck结构和功能特点
1.p56lck的结构 p56lck是由lck基因编码的分子量为56kDa的单链分子,由509个氨基酸残基组成。在小鼠,lck基因定位于4号染色体,人lck基因定位于1号染色体的1p32-35区间。同其它具有PTK活性的生长因子受体如EGF-R和PDGF-R等比较,p56lck分了属于非受体型蛋白酪氨酸激酶,无胞膜外区,其N-端通过豆蔻酸(myristic acid)与细胞膜内侧面相连。p56lck分子结构可以分为四个功能区:(1)膜接触区,位于N-端,N-端第2位Gly能结合豆蔻酸,通过豆蔻酸与细胞膜内侧面相连;(2)底物作用区,该区的氨基酸结构不同于src家族的大部分其它成员;(3)催化区或激酶区,这一区域在氨基酸组成上与其它src家族成员高度同源;(4)调节区,位于羧基端,可能与p56lck的PTK活性的特异调节有关(图8-6)。

图8-6 p56lck分子的功能区
2.p56lck的功能特点 p56lck底物作用区存在几个位置尚不明确的Ser磷酸化位点;催化区Lys273是ATP结合点,Tyr394为自身磷酸化位点;调节区Tyr505是调性磷酸化位点。在静止细胞中,p56lck在Tyr505上发生磷酸化,但当细胞活化时这个残基则发生去磷酸化,随之发生激酶的活化以及Tyr394的自身磷酸化。目前研究认为,CD45通过使Tyr505去磷酸化对p56lck起正调控作用;而p50csk是使Tyr505发生磷酸化对p56lck则起负调控作用。CD45和p50csk对其它src家族PTKs也发挥着同样的调节作用。
(二)p56lck与CD4/CD8分子
1.p56lck的分布 CD4和CD8以共受体形式(coreceptor)表达于成熟的T淋巴细胞,尽管CD4和CD8跨膜分子都属于免疫球蛋白超家族,但它们之间仅存在有限的同源性。CD4和CD8分子分别与MHCⅡ类和MHc I类分子的恒定区决定簇相互作用。CD4是分子量为55-60kDa的糖蛋白,以单体形式表达。CD8是由二致辞体组成,有两种形式:一种是由两条α链(32-34kDa)组成的同源二聚体;另一种形式是由α链和β链(25-26kDa)组成的异源二聚体。CD4和CD8分子中胞浆内部分子不具有激酶功能区,因此它们与受体酪氨酸激酶如EGF-R或PDGF-R无同源性。目前已经证实CD4和CD8均与信号转导成份p56lckPTK相连接。p56lck几乎在所有淋巴细胞中表达,包括全部成熟T细胞和胸腺细胞,提示它可能在T细胞活化和分化调节过程中起作用。有关p56lck在T细胞活化信号中正调节作用的最初发现是p56lck直接同CD4或CD8复合受体分子胞浆内功能域相连接,在体外具有酪氨酸激酶活性,在体内,T细胞受到刺激后酪氨酸磷酸化水平增加。即使在静止的鼠CD4-T细胞中也有大约50%细胞内p56lck是同CD4糖蛋白相连接。
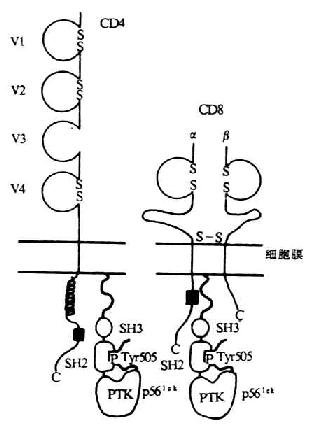
图8-7 p56lck同CD4、CD8作用的模式图
2.p56lck与CD4/CD8分子的连接 p56lck同CD4/CD8相互作用的区域位于分子的氨基端的底物作用区(图8-7),此区域在src相关PTKs中是独特的,含有4个半胱氨酸,这对于p56lck与CD4和CD8相互作用是必须的。底物作用区中6个带负电荷的氨基酸可使p56lck与CD4/CD8结合位点内相对应的碱性氨基酸残基相结合。在CD4和CD8α分子的胞浆内有一个含13个氨基酸的相似区域,经突变研究和肽竞争分析法证实此区域可能为p56lck结合区域。CD4和CD8α含有两个对于p56lck结合起关键作用的半胱氨酸以及与p56lck氨基末端负电荷相互作用的5个带正电荷的氨基酸。证实此区域中有6个氨基酸直接与p56lck结合有关,如果把这6个氨基酸残基从CD8α胞浆功能域转移到一个非相关蛋白水泡性口炎病毒糖蛋白(VSV-G)胞浆功能区域上,p56lck即可结合到这个杂交分子上去。
3.p56lck参与T细胞活化 用蛋白激酶C激活剂刺激T淋巴细胞可引起p56lck从CD4分子上解离下来,随后发生CD4内化。CD8分子在这方面不同于CD4,CD8+T淋巴细胞经PKC激活剂刺激事对CD8分子内化以及CD8-p56lck的解离影响极微。现已提示,p56lck可能以生长因子PTK受体相同方式起作用,即TCR结合与MHC相连的抗原后,CD4中CD8与MHC相互作用,与CD4或CD8相连接的p56lck补充到TCR多肽胞浆内部分靠得很近的区域,然后发生PTK的活化以及TCR内底物(?)和/或其邻近分子(PLCγ1?)的磷酸化。p56lck除参与抗原诱导的淋巴细胞活化外,还可能参与T细胞的分化和成熟。在T细胞分化的所有阶段,包括CD4、CD8双阳性胸腺细胞都可检测到p56lck与CD4和CD8形成的复合物。
(三)p56lck与非CD4/CD8分子
p56lck除了同CD4和CD8形成复合体外,还可能同另外一些参与信号转导的细胞受体分子形成稳定的复合物。
1.p56lck与IL-2R的关系 IL-2R由α、β、γ三条肽链组成。三条肽的不同组合即αβγ,βγ以及单独α分别构成IL-2的高、中、低三种亲和力的受体。由于IL-2Rα链在胞浆内仅有一个较短的功能域(13个氨基酸残基),所以介导IL-2R信号转导主要为β和γ链。IL-2Rβ链胞浆内有两个结构域:其中一个靠近细胞膜,富含丝氨酸,对于IL-2诱导的增殖信号具有重要作用;另一结构域远离细胞膜,富含酸性氨基酸,为酪氨酸激酶物理连接部位。经IL-2Rβ链免疫沉淀后进行免疫印迹也证实了IL-2Rβ链同p56lck相连。体外系统中也发现IL-2结合到IL-2R后可促进p56lck活性,引起IL-2Rβ链的酪氨酸磷酸化。IL-2Rγ链胞浆内含86个氨基酸,无激酶功能区,但具有一个与SH-2同源的区域,可能参与IL-2R介导的信号转导。最近研究表明,IL-2Rγ链为IL-4、IL-7、IL-9、IL-13等细胞因子受体所共用(common chain,γC)。
2.p56lck与其它分子的关系 同p56lck相互作用的其它一组分子有人T细胞糖磷脂酰肌醇(glycophosphatidylinositol,GPI)连接的蛋白包括CD59、CD55、CD48以及小鼠T细胞中的Thy-1。应用免疫沉淀方法均能使这些分子同p56lck发生共免疫沉淀。此外,将T淋巴细胞同针对GPI连接的膜分子特异性抗体孵育,然后加入二抗使其交联,均发生几种内源性底物的酪氨酸磷酸化。
许多GPI连接的细胞表面分子可能以两种不同的形式表达在T细胞中,这是由相应分子mRNA不同拼接所致。一种形式是通过GPI附着在细胞膜上,另一种形式具有跨膜区和胞浆的PLC去掉所有GPI相连的膜蛋白后进行免疫沉淀,并分析它们相连的激酶活性,发现大多数酪氨酸磷酸化活性已丧失,表明GPI连接物对于同p56lck相结合是必需的。
采用烷基化试剂或金属离子结合试剂进一步证实了CD4/CD8多肽和GPI连接膜蛋白是分别通过两种不同的机理与p56lck相互作用的。这些试剂可以干扰p56lck同CD4或CD8分子的相互作用,因为p56lck同CD4或CD8的相互作用依赖游离半胱氨酸的存在,并需要金属离子使之稳定这种结合;而p56lck与GPI相连结的蛋白的结合则不受这些试剂影响。这些结果表明,p56lck通过两种或更多不同相互作用机理,直接或间接地同多种细胞表面蛋白形成复合物。