二、T淋巴细胞亚群
应用CD4和CD8单克隆抗体可将外周淋巴器官或外周血中的T细胞分为CD4+CD8-和CD4-CD8+两个主要的亚群。每个亚群按照某些表面标志和功能又可分为不同的功能亚群。
(一)CD4阳性细胞群
1.Th1和Th2亚群 应用Th细胞克隆培养技术和细胞因子产生的不同,已发现小鼠CD4阳性细胞群是一个不均一的亚群,可分为Th1和Th2,主要区别见表7-6。
Th1细胞能合成IL-2、IFN-γ、,LT、IL-3、TNF-α和GM-CSF,但不能合成IL-4、IIL-5、IL-6、IL-10和IL-13;而Th2能合成TNF-α、IL-3、GM-CSF、IL-4、IL-5、IL-6、IIL-10(细胞因子合成抑制因子,CSIF)和IL-13,不能合成IL-2、IFN-γ和LT。此外Th1和Th2都能分泌三巨噬细胞炎症蛋白和前脑啡肽原。Th1和Th2都能辅助B合成抗体,但辅助的强度和性质不同。体外实验表明,IL-4明显促进B细胞合成和分泌IgE,如使LPS刺激小鼠B细胞合成IgE能力增强10-100倍。少量IFN-γ能完全阴断IL-4对IgE合成的促进作用。Th2分泌IL-4对IgE合成有正调节作用,而Th1分泌IFN-γ则起负调节作用。此外,Th2通过分泌IL-4和IL-5辅助IgA合成,分泌IL-10(CSIF),抑制Th1细胞合成细胞因子,而Th1对IgG1合成则有抑制作用,但辅助其它几种类型Ig的合成。由于Th1和Th2合成淋巴因子的种类不同,因而介导不同的超敏反应。IL-3和IL-4均能促进肥大细胞增殖,且相互有协同作用,IL-5除辅助B细胞合成IgA外,还能刺激骨髓嗜酸性粒细胞的集落形成,因而Th2与速发型超敏反应关系密切。Th1通过产生IFN-γ阻断IgE合成,对速发型超敏反应有抑制作用。Th1与迟发型超敏反应有关,可能与IL-2、IFN-γ等对巨噬细胞活化和促进CTL分化作用有关,此外LT也有直接杀伤靶细胞作用。两群Th克隆均能诱导抗原提呈细胞(APC)表达MHCⅡ类抗原,Th1通过IFN-γ诱导Mφ表达Ia抗原,而Th2通过IL-4对Mφ和B细胞Ia抗原表达起正调节作用。在人类Th1和Th2细胞亚群尚未得到最后证实。从目前发表资料来看,CD4+CD45RO+前体细胞向Th2效应细胞分化,而IFN-γ则对前体细胞向Th2分化过程起抑制作用,因此IL-4和IFN-γ在决定CD4+CD45RO+前体细胞向Th1或Th2分化过程中起着重要的调节作用。人T细胞经多克隆活化后,在CD4阳性细胞中IL-4mRNA阳性比便不到5%,而60%的CD4+细胞有IFN-γ和IL-2mRNA的转录。
表7-6 小鼠Th1和Th2亚群的比较
| Th1 | Th2 | ||
| 合成淋巴因子种类 | IL-2 | + | - |
| IFN-γ | + | - | |
| LT | + | - | |
| IL-3 | + | + | |
| TNF-α | + | + | |
| GM-CSF | + | + | |
| IL-4 | - | + | |
| IL-5 | - | + | |
| IL-6 | - | + | |
| IL-10 | - | + | |
| IL-13 | - | + | |
| 辅助B细胞 | + | ++ | |
| 辅助IgE合成 | - | + | |
| 促进肥大细胞增殖 | - | + | |
| 介导超敏反应类型 | 迟发型 | 速发型 | |
| 促进Ia表达细胞 | Mφ | B,Mφ |
介导迟发型超敏反应(delayed type hypersensitivity,DTH)的T细胞亚群称为TDTH,表面标志CD3+CD4+CD8-,可能相当于小鼠的Th1亚群。当曾被变应原致敏的TDTH再次与变应原相遇后,可释放出多种细胞因子,参与迟发型(Ⅳ型)超敏反应的发生。
表7-7 TDTH释放细胞因子参与迟发型超敏反应
| 作用对象 | 细 胞 因 子 | 生物学活性 |
| 巨噬细胞 | 巨噬细胞趋化因子(MCF) | 招引、聚集、活化巨噬细胞, |
| 巨噬细胞移动抑制因子(MIF) | 增强巨噬细胞吞噬和杀伤功能 | |
| 巨噬细胞活化因子(MAF) | ||
| IFN-γ | ||
| 白细胞 | 白细胞移动抑制因子(LIF) | 招引、聚集白细胞 |
| 淋巴细胞 | IL-2 | 淋巴细胞局部浸润、增殖 |
| IFN-γ | 增强杀伤功能 | |
| 皮肤血管 | 皮肤反应因子(SRF) | 增加血管通透性 |
| 靶细胞 | 淋巴毒素(LT) | 杀伤靶细胞 |
MCF: macrophage chemotactic factor
MIF: macrophage migration inhibition factor
MAF: macrophage activating factor
LIF: leukocyte migration inhibitory factor
SRF: skin reactive factor
LT: lymphotoxin
2.抑制细胞诱导亚群和辅助细胞诱导亚群 应用CD45RA、CD45RO、CD29和CD31单克隆抗体可将CD4阳性细胞群分为抑制细胞诱导亚群和辅助细胞诱导亚群。两个亚群的表面标志和功能比较见表7-8。
(1)CD31:最近发现CD31是一种新的、激活后表达水平不发生明显变化的抑制细胞诱导亚群的表面标记。CD31是一种血小板-内皮细胞胞粘附分子(PECAM,gpⅡa),分子量为140kDa,其结构属于免疫球蛋白超家族成员。从外周血新鲜分离的CD4细胞中,CD31McAb主要与CD45RA亚群反应,对B细胞合成IgG辅助作用不明显,对ConA和自身MHC(自身MLR)反应较为敏感;而CD31-的CD4细胞群中,发现有大量辅助B细胞合成IgG的活性和对某些抗原刺激的回忆反应。CD45RA+的CD4细胞大激活后,尽管细胞表面丢失CD45RA,但表面CD31的表达仍不发生明显变化;而CD45RO+CD45RA-的CD4细胞激活后不能获得CD31表达。由于CD31在CD4细胞激活后仍不变化,对于鉴别抑制细胞诱导亚群和辅助细胞诱导亚群是一种有用的标志。
迄今为止,许多粘附分子如 CD11a/CD18(LFA-1),LFA-3,CD2和CD29(VLAβ链)主要表达在CD45RO+T细胞表面。而CD31则表达在CD45RA+CD4细胞表面。抗CD31McAb作用于naive T细胞能触发其VLA-4介导的粘附作用。内皮细胞表面CD31及其配体与T细胞表面CD31及其配体相互作用很可能触发整合素介导的粘附作用。CD31如何参与CD45RA+CD4+T细胞功能以及诱导抑制性T细胞产生还有待进一步研究。
(2)CD45:CD45为异构型分子。CD45细胞膜外部多肽链可由A、B和C三种外显子编码。人幼稚T细胞只表达被抗CD45RA识别的CD45A型;记忆T细胞不表达任何A、B、C外显子产物而被抗CD45RO识别。抗CD45RA和抗CD45RO识别的都是休止型细胞,抗CD45RO所识别的记忆T细胞往往也可低水平表达一系列活化表面标记,如CD25、MHCⅡ类抗原、CD54、CD26等,提示这类细胞可能新近被激活过,由此推论记忆T细胞可能是由于持久性抗原或交叉抗原的低剂量、持续刺激得以维持其长时间存活。体外实验观察到细胞活化后见有从CD45RA向CD45RO的单向性转变,这与幼稚T细胞向记忆T细胞分化相平行。
表7-8 抑制细胞诱导亚群和辅助细胞诱导亚群的比较
| 抑制细胞诱导亚群 | 辅助细胞诱导亚群 | |
| 表 型 CD4 | + | + |
| CD29 | 低密度 | 高密度 |
| CD31 | + | - |
| CD45 | CD45RA | CD45RO |
| 粘附分子(LFA、) | + | +++ |
| 增殖功能 | + | +++ |
| CD3McAb | + | +++ |
| +++ | + | |
| 抗原刺激再次反应 | - | +++ |
| 同种异体抗原 | +++ | +++ |
| 自身MLC | +++ | + |
| 诱导和辅助功能 | ||
| 辅助B细胞产生抗体 | ± | +++ |
| 诱导Ts | +++ | - |
| 诱导细胞介导的淋巴细胞溶解作用(CML) | ± | +++ |
| 细胞发育阶段 | 天然(naive) | 记忆(memory) |
(3)自身混合淋巴细胞反应:外周血B细胞和单核细胞等非T细胞在体外培养时能诱导某些自身T细胞发生增殖反应,称为自身混合淋巴细胞的应(autologous mixed lymphocyte reaction ,AMLR)。这一部分T细胞称为自身反应性T细胞。作为刺激细胞的B细胞和单核细胞主要是通过其细胞表面的MHCⅡ类抗原来刺激自身反应性T细胞,在体外培养时加入抗MHCⅡ类抗原的抗体可阻断AMLR。关玩弄AMLR的生理意义尚不清楚,可能是机体的一种免疫调节机制。
(2)CD8阳性细胞群
根据CD28阳性或阴性可将CD8+细胞分为细胞毒性T细胞(CD8+CD28+)和抑制性T细胞(CD28+CD28-)。CD28McAb能与60-80%T细胞发生反应,包括全部CD4细胞和部分CD8细胞。
1.Tc(CTL) 在人类CTL表型为CD3+CD4-8+CD28+。小鼠CTL表型为Thy-1+、Lyt-1+、Lyt-2+/Lyt-3+。
(1)CTL的分化:静止的CTL以前体细胞(precursor)(CTL-P)形式存在,外来抗原进入机体被抗原提呈细胞(APC)加工处理,形成外来抗原与APC自身MHc I类抗原的复合物,被相应CTL克隆细胞膜表面TCR/CD3所识别,抗原刺激信号和APC释放IL-1共同存在的条件下,CTL-p被活化,并表达IL-2R、IL-4R、IL-6R等多种细胞因子受体,在IL-2、IL-4、IL-6、IFN-γ等细胞因子诱导下,迅速增殖,并分化为成熟的效应杀伤性T细胞(effector CTL)。CTL具有识别抗原的特异性,即能杀伤具有特定的外来抗原(如病毒感染靶细胞膜表面的病毒抗原)与自身MHc I类抗原结合的复合物的靶细胞。有关CTL杀伤靶细胞受到MHC I类抗原的限制,参见第五章“主要组织相容性复合体”第四节。从肿瘤组织周围分离获得的CTL称为肿瘤浸润淋巴细胞(tumor infiltrating lymphocyte , TIL)。TIL在体外加IL-2培养后,具有很高的杀伤肿瘤作用,目前已用于临床的肿瘤治疗。
(2)CTL的识别机制:多种粘附分子参与CTL对靶细胞的识别和粘附,主要有:①LFA-1/ICAM-1、ICAM-2、ICAM-3,可溶性ICAM-1(sICAM-1)可抑制CTL杀伤肿瘤细胞;②CD2/LFA-3(CD58),抗CD2McAb或抗CD58 McAb均可抑制CTL效应细胞对靶细胞的杀伤;③CD8/MHc I类抗原的非多态性结构域。
(3)CTL的杀伤机制:TCL杀伤靶细胞的机理目前认为主要通过释放多种的介质和因子介导的。
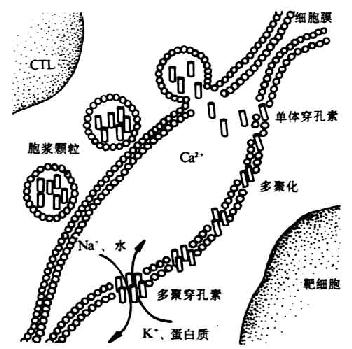
①穿孔素(perforin):又称成孔蛋白(pore-foming protein,PFP)、C9相关蛋白(C9 related protein)或溶细胞素(cytolysin),贮存于电子稠密胞浆颗粒(electron-densecytoplasmic granules),成熟的穿孔素分子由534个氨基酸残基组成,分子量为56-75kDa,IP为6.4,穿孔素分子中央部位170-390之间的氨基酸序列与C9328-560氨酸酸序列约有20%同源性,这个区域与穿孔素和C9的多聚化和以管状形式插入到细胞膜有关。在杀伤相时,CTL细胞脱颗粒,穿孔素从颗粒中释放,在Ca2+存在下,插入靶细胞膜上,并多聚化形成管状的多聚穿孔素(polyperforin),约含12-16个穿孔素分子,分子量可达1000kDa。多聚穿孔素在靶细胞膜上形成穿膜的管状结构,内径平均16nm。这种异常的通道使Na+、水分进入靶细胞内,K+及大分子物质(如蛋白质)从靶细胞内流出,改变细胞渗透压,最终导致细胞溶解。此过程与补体介导的溶细胞过程类似,溶解细胞过程比较迅速。CTL本身可能释放A型硫酸软骨素蛋白聚糖(proteoglycans of chondroitinsulphate A type)、硫酸软骨素A限制因子(homologous restriction factor,HRF),因此可避免穿孔素对CTL自身细胞的攻击。
图7-3 CTL释放穿孔素杀伤靶细胞机理示意图
②丝氨酸酯酶(serine estersse):活化CTL释放多种丝氨酸酯酶,如CTLA-1(又称CCP1或granzyme B)、CTLA-3(又称H因子或granzyme A),其作用可能类似补体激活过程中的酯酶作用,通过活化穿孔素而促进杀伤作用。
③淋巴毒素(lymphotoxin,LT):又称肿瘤坏死因子-β(TNF-β),LT可直接杀伤靶细胞,但杀伤过程较慢,其杀伤机理参见第四章“细胞因子及其受体”第二节中肿瘤坏死因子。
2.Ts和Ts亚群 抑制性T细胞(suppressor T lymphocyte,Ts)对免疫应答有重要的负调节功能,抑制性T细胞功能的异常,常与T自身免疫性疾病、第I型超敏反应等疾病发生有关。
(1)抑制性T细胞的证实:绵羊红细胞(sheep red blood cell , SRBC)对于小鼠是良好的免疫原,合适剂量的SRBC可诱导小鼠产生高效价的抗SRBC抗体。当过高剂量SRBC免疫小鼠时,则抗体合成水平反而明显下降,称为高剂量免疫耐受。动物实验研究发现,将高剂量免疫耐受小鼠脾细胞转移到免疫原剂量刺激的小鼠体内时,则小鼠抗体应答水平明显下降。如高剂量免疫耐受小鼠脾细胞经抗Thy-1和补体处理后再转移到免疫原剂量免疫的小鼠体内,则高剂量免疫耐受小鼠脾细胞的抑制作用消失。实验证明了在高剂量免疫耐受小鼠的脾细胞中存在有抑制作用的T细胞。
表7-9 抑制性T细胞的证实
| (1) | 免疫原剂量---→抗体应答+++ |
| SRBS免疫小鼠 | |
| (2)高剂量SRBC---------------------------------→抗体应答+ | |
| 免疫耐受小鼠 | |
| (3)高剂量免疫-→转移未处理脾细胞-→免疫原剂量-→抗体应答+ | |
| 耐受小鼠 免疫小鼠 | |
| (4)高剂量免疫-→转移抗Thy-1+ ----→免疫原剂量-→抗体应答+++ | |
| 耐受小鼠 补体处理脾细胞 免疫小鼠 | |
进一步研究证明,这种抑制细胞的表型为CD3+CD4-CD8+(小鼠CD8单抗常用Lyt-2)。人的抑制性T细胞表型为CD3+CD4-CD8+CD28-。Ts细胞不仅对B细胞合成和分泌抗体有抑制作用,而且对Th辅助作用、迟发型超敏反应以及Tc介导的细胞毒作用都有负调节作用。
(2)Ts细胞的亚群:Ts细胞还可分为Ts1、Ts2和Ts3不同亚群,分别起着诱导抑制、转导抑制和发挥抑制效应的作用。它们之间相互作用的确切机理还不十分清楚,可能是通过释放可溶性介质相互作用的。Ts1(Tsi,抗原特异性抑制性T细胞)分泌TsF1(TsiF,抑制诱导因子)→作用于Ts2(Tst,抑制转导细胞),分泌TsF2(Tst F)→作用于Ts3(Tse,抑制效应细胞),分泌Ts3F(TseF),作用于Th细胞,通过对Th的抑制作用,从而对各种免疫功能起负调节作用。Ts细胞群具有高度异质性,除Ts1、Ts2、Ts3亚群外,还有一群反抑制性T细胞亚群(contra-suppressor T cel,Tcsl)。Tcs活化后分泌反抑制性T细胞因子TcsF,直接作用于Th细胞,解除Ts细胞的抑制作用,使Th细胞恢复辅助活性。