�ڶ��ڡ�ϸ�����ӵĽṹ������ѧ����
�������ڻ��̼�����Ѹ�ٷ�չ������ϸ�����ӵ�cDNA��¡��óɹ���������˸��Ի�����������ϸ�����ӵ�ϸ������ѧ�ͷ�������ѧ���о��ṩ�˱�Ҫ��ǰ�ᡣĿǰ����IL-2��EPO��G-CSF��GM-CSF��IFN-�ú�IFN-����ϸ������Ͷ���г����и����ϸ���������ڽ��в�ͬ�ڵ��ٴ���֤��ϸ����������ʽͶ���г���Ϊ��Ʒǰ�����뾭���ٴ�ǰɸѡʵ�顢�ٴ����ڡ����ڡ��������õȼ����ε���֤��
����һ����ϸ�����أ�IL��
������1979��ڶ�������ܰ�����ר�����ۻ��ϣ������Ե���-����ϸ����T�ܰ�ϸ�������ڵ�ijЩ�������Է������ߵ��ں�����֢��Ӧ�������õ����ӳ�Ϊ��ϸ�����أ�interleukin��IL����Ŀǰ��֪����IL�����Ե���-����ϸ�����ܰ�ϸ�����������ϸ��������ʽ�����İ�ϸ��������IL-1��IL-15��
������һ��IL-1
����IL-1��һ�ֵ������ӡ�1972��Gery�ȷ����˰�ϸ�������������к���һ�ֿ��������ʣ��������ʿɴٽ�С������ϸ����ֲ��Ѫ���أ�PHA������˿���ѷ�Ӧ���������Ϊ�ܰ�ϸ���������ӣ�lymphocyte-activating factor,LAF)����Դ����ԭ�ʣ�endogenous pyrogen)���ƹ�ϸ���������ӣ�osteoclast activating factor)��������ϸ�������������ӣ�melanoma growth inhibitory factor)�ȣ�1979�����ͳһ����ΪIL-1��
����1.IL-1�IJ�����IL-1���ɶ���ϸ���ϳɺͷ��ڡ�
������1������ϸ��������ϸ�����縹ǻճ��ϸ��peritoneal cell,PC)����ͻ״ϸ��������ȡ��ԭ���帴�������ڿ�ԭ��ʹ����пɲ���IL-1���ڴ�����̼����̼�����Ѫ������ϸ����PBMC)�����£�IL-1��mRNAˮƽҪ����IL-1��mRNa 20��25����
������2��С�����ϸ��ϸ��ϵP388D1��J774��PU5-1.8��WEHI-3�Լ���ǰ����ϸ����U937����֬���ǣ�LPS���̼����ܷ��ڴ�����IL-1��
������3����Ƥϸ����NKϸ����Bϸ��������άϸ������Ƥϸ�����Խ�����״ϸ������С��ϵĤϸ����mesangial cell������Ĥ����ϸ����synovial lining cell����ƽ����ϸ������Ƥϸ����̥��ϸ������Ѫ��ϸ����PMN����ijЩ��������ɲ���IL-1��
�����������ؿ���ֱ��Ӱ�쵥��-����ϸ��IL-1�ķ��ڣ���ϸ�����������ϸ���������ӣ�MAF��������̼����ӣ�CSF����IFN-����IFN-�ö�IL-1����������ǿ���á���LPS��PPD��BCG����˹�ؾ������������������ⶾ�أ������Ҳ��IL-1�����Ĵ̼�����������Ѫ��20pg/ml�ڶ��ش̼�����ϸ�����ɲ���IL-1�������һ�����������£��������ڶ��ص���Ⱦ�ɴ̼�����ϸ������IL-1��10ng/mlLPS��ʹ����ϸ���ϳ�IL-1����100�����۵���øC��PKC���ļ���������ⶹޢ����phorbol myristate acetate,PMA)�Լ����������壨calcium ionophore)��A23187������ǿ�ҵĴ̼����á���Ƥ����̴���ǰ���������IL-1�IJ������������á�
����2.IL-1�ķ��ӽṹ�ͻ�����������IL-1����IL-1�»�����ֱ�Ϊ10.5kb��7.8kb���˺�С��IL-1����λ��2��Ⱦɫ�壬����7�������ӡ�IL-1ǰ��(ProIL-1)Ϊ31kDa��ͨ������ˮ��ø�ѽ��γɳ����IL-1���ӡ�IL-1�ڲ�ͬ�������нϸ�ͬԴ�ԡ��ڰ�����ˮƽ�ϣ�IL-1����IL-1���ڲ�ͬ����ͬԴ�Էֱ�Ϊ60%��70%��75%��78%������ͬһ������IL-1����IL-1��ͬԴ��ֻ��25%����IL-1����PI5.0����IL-1�£�P17.0���ֱ���159��153��������л���ɣ�������Լ17.5kDa��ͬԴ��Ϊ28%��IL-1���ӵ���ø���С���ͬϸ��������IL-1�ĵȵ����������졣
����3.IL-1�����塡Tϸ��������άϸ������IL-1����Ϊ80kDa����Bϸ����Ϊ68kDa������������IL-1�����Dz�ͬ������P80IL-1R��ΪIL-1RtI��CDw121a),P68IL-IR��ΪIL-1Rt��CDw121b)��
��4-1��IL-1 Rt ���IL-1Rt��ıȽ�
| IL-1 Rt�� | IL-1Rt�� | |
| �ṹ | ||
| ͬԴ�� | Ig�����壨3��C2���� | Ig�����壨3��C2���� |
| ��������kDa) | 80 | 68 |
| ��������Ŀ | 549 | 384 |
| ��Ĥ�� | 319 | 329 |
| ����Ĥ | 20 | 26 |
| ������ | 213 | 29 |
| ��Ҫ�ֲ�ϸ�� | �㷺��Tϸ��������άϸ���� | EBVת��Bϸ����Raji��PMN������ϸ���� |
| �������� | ���� | ���� |
| ������� | IL-1��>IL-1�� | IL-1��>IL-1�� |
| ��˿����/�հ��� | +���ź�ת���� | �������в��壩 |
| ���л����ữ | �� | �� |
| ���������Ϻ�仯 | ���ڻ� | ���� |
������1��IL-1Rt��IL-1Rt�� cDNA��¡���˺�����ѻ�óɹ���IL-1Rt��Ϊ��Ĥ���ף���Ĥ������3���ṹ���������׳����壬��Ĥ����20��������л�������������˿������հ���л�����IL-1��IL-1Rt���Ϻ�˿������հ���ܿ챻���ữ��ͨ������תȾHelaϸ��ʵ��֤����IL-1Rt��N��2���ṹ�����������йء����N��17��������Ƭ�ε�McAb�����IL-1Rt����IL-1��ϡ�IL-1��IL-Rt���Ϻ����ڻ���internalization)������άϸ����ƽ����ϸ����Ҫ����IL-1Rt��һ����˵��IL-1Rt�����IL-1����IL-1�����ϣ���IL-1������������������ϸߣ���IL-1������������Ͻϸߡ�IL-1�벻ͬ������ͬϸ����Ϻ������ѧЧӦ����������˺���IL-1����ϵ�����Ƥϸ���ϵ�������ͬ��������������ѧЧӦ����ȫ��ͬ����IL-1Rt��McAb�����ں������������IL-1������ѧЧӦ��
������2��IL-1Rt����Ҫ�ֲ���EBVת����Bϸ����Rajiϸ��������ϸ����̥�̡�Th2��¡���Tϸ����PMN����ϸ���ȡ���Ĥ������3���ṹ���������׳����壬��IL-1Rt��֮����28%������ͬԴ�ԣ���Ĥ���и��ߵ�ͬԴ�ԣ���������Ҫ�Ȣ�������̣������ڽ鵼�źŴ������в��IL-1��IL-1Rt���Ϻ��������������IL-1Rt�����������ڻ���IL-1Rt����ˮ��øˮ�����γɿ����Ե�IL-1��ϵ��ף�soluble IL-1 binding protein,sIL-1BP)��46kDa����IL-1���нϸ�����������Ȼ�����sIL-1BP��IL-1�µ����Ƽ���������IL-1R��soluble IL-1 receptor,sIL-1R)����Ч��ֹС��������ֲ�ųⷴӦ������Lewis�����ʵ���Թؽ������Դ����ס�
������Tϸ���У�CD4����ϸ����ȺIL-1�������Ҫ����CD8����Tϸ����Ⱥ����С��������ϸ��ϵEL-4Ϊģ�ͣ�����IL-1����IL-1�¿ɽ�ϵ���ͬ�ĸ��������塣���ѷ�����������ȱ�ݲ��˵�Tϸ������IL-1Rȱ�ݣ��Կ�ԭ�̼���������ֳ��Ӧ��Ҳ������IL-2�������������²�����Ⱦ��
����4.IL-1���������ijЩ��Ѫ������Ѫ��������Լ�����ϸ�����������з���һ�ֶ������ʵ�IL-1�������������ӣ���ΪIL-1�������interleukin 1 receptor antagonist,IL-1ra),�ֳ�IL-1���������ף�IL-1 receptor antagonist protein, IRAP)��
������1��IL-1ra�IJ�����IL-1ra�������LPS�̼��ĵ���ϸ����PMA��PHA��CSF�̼��ĵ���ϸ��ϵ������
������2��IL-1ra���ӵĽṹ�ͻ���IL-ra�����¡1990���óɹ���������IL-1����IL-1�º�IL-1ra����λ��2��Ⱦɫ�塣IL-1ra cDNA����Ķ���Ϊ17kDa���ǻ����������Ϊ25kDa�����ǻ���IL-1ra���Բ��DZ��衣δ�����IL-1ra����Ϊ177��������л���������N��25���������Ϊ��ˮ�����ᣬ���ɵ��͵��ź���˳���������152��������л���ɡ���65��68��116��122λ����4�����صİ��װ���л���Cys65-116��Cys68-122���γ����ڶ��������cDNA����İ���������IL-1ra��IL-1����IL-1�·ֱ���19%��26%��ͬԴ�ԣ��˺�С���IL-1ra��77%ͬԴ�ԡ�
������3��IL-1ra������ѧ���ã�IL-1ra�������Ե�����Tϸ������IL-1R��IL-1��ϣ���������TNF��IL-2����Ӧ����Ľ�ϡ�IL-1ra����IL-1ֱ�ӽ�ϣ�����һ��IL-1��IL-1R���ϵľ����������rIL-1ra����ͺ͢���IL-1R���ܽ�ϣ�����IL-1Rt���ϵ�����Ҫ������IL-1Rt���ϵ�������rIL-1ra��rIL-1����rIL-1����IL-1Rt���ϵ������Ƚ��������rIL-1ra��rIL-1����IL-1Rt���ϵ�����Ҫ����rIL-1����IL-1Rt���ϵ�������IL-1ra������IL-1�̼���Ĥϸ��PGE2�IJ���������ϸ����ԭø�ϳɣ���������ϸ������ֳ�Լ�������ϸ������������ϸ������Ƥϸ����ճ���������ڿ�����IL-1����ķ��ȡ�IL-1ra�ɽ��Tϸ���ͳ���άϸ������IL-1Rt��,Ҳ������IL-1��PMN��Bϸ������������ϸ����Ѫ��ϸ��IL-1Rt��Ľ�ϡ�IL-1ra������PBMC������ϸ�������������ܰ�ϸ����Ѫ��ϸ���Է���ֳ���Է�����IL-1��IL-6��GM-CSF�������ڣ�IL-1ra����ֹLPS����ļ����������������߸��������յ�����֢������С�������ֲ��GVHR�ķ�������ߴ���ʣ����⣬���ɷ��ζ���ʵ���������Խ᳦�ס�
������4��IL-1ra���ٴ���������Ѫ��IL-1raˮƽ��200pg/ml���£���Ⱦ����֢�Լ��ڶ���Ѫ֢����Ѫ����IL-1raˮƽ�����ߵ�8ng/ml��Ӧ��IL-1ra���ư�Ѫ֢�ѽ�������ٴ���֤�������������½�������IL-1ra�������ʪ�Թؽ���Ҳ�ѿ�ʼ���ٴ���֤��IL-1ra�����ڵĸ����ú�С����Ӧ�ü����ϴ����IL��1��50%����ѧЧӦʱ����IL-1ra��������IL-1������10��500����
����5.IL-1������ѧ���á�IL-1���й㷺�����ߵ������ã��������Ⱥͽ鵼��֢�����ã���������ѧ������ͨ������Ӧ�����������϶��鵼�ģ�IL-1���������µ�Ũ�Ƚ���10-12��10-14M֮�䡣IL-1���������Ե����������ԣ���IL-1��������С��Դ�Ե�ϸ������Ҫ����IL-1Rt���ϸ���ƺ��ȱ���IL-1Rt��ϸ����������������ԡ�
������1���ٽ�����ϸ����Tϸ���Ļ����ֳ�ͷֻ���Tϸ������ԭ����˿����ԭ��TCR/CD3�̼������IL-1���壬��IL-1������Tϸ���������G0�ڽ���G1�ڡ�����Tϸ������IL-2��IFN-�á�GM-CSF��IL-4��ϸ�����ӣ�������IL-2�������T������ֳ�ͷֻ���IL-1��������Tϸ������MHC���ԭ�ı��IL-1���յ�ɱ����T�ܰ�ϸ����CTL���ķֻ����ڻ���ܰ�ϸ��������MLC���У�IL-1�յ�CTL�IJ���������ͨ���ٽ�Tϸ������IL-2��IFN-�á�IL-6��ЭͬIL-1�Tϸ���ʹ̼�IL-2�IJ�����
������2���ٽ�Bϸ�����ܣ�ЭͬIL-4��ϸ�����Ӵ̼�Bϸ������ֳ�ͷֻ����ٽ������ĺϳɺͷ��ڣ��������ÿ�����ͨ��IL-1�յ�PBMC����IL-6���鵼�ġ�
������3���̼�������ܸ�ϸ������ֳ������֤ʵIL-1��Stanley(1986)������Ѫϸ��������-1��hemopoietin-1)��ͬһ�ַ��ӡ�IL-1�̼���Ѫϸ���ͳ���άϸ������CSF��������Ѫϸ��CSF�������������ЭͬIL-3��IL-6��G-CSF��M-CSF��GM-CSF��SCF�����Ӵ̼���Ѫ���ܣ�������ϵ��ϸ���;�ϵ��ϸ�����д̼����á����⣬IL-1�ɴ̼���ϸ������SCF��IL-1�������������ڸ�ϸ���������ϸ����Go�ڽ�����ֳ���ڡ�IL-1��Ԥ��������ɵĹ��������ѽ����ٴ�������֤��
������4����ǿNKϸ����ɱ�˻��ԣ�ͨ�����NKϸ����IL-2��ϸ�����ӵ���������ǿ��ɱ�˻��ԣ�IL-1��IL-2��IFN��Эͬ�̼�NKϸ�����Ե����á�
������5���ٽ��������߷��ӵĻ�������IL-1��IL-2��IL-3��IL-4��IL-5��IL-6��IL-7��IL-8��TNF-����TNF-�¡�INF-�¡�G-CSF��GM-CSF��GM-CSF��IL-2R������Tac)������C2��Bf��ճ�������Լ�c-fos��c-myc��c-jun��ԭ������ı��C-fos��C-jun��ɻ����-1��AP-1�����IL-2����������ӣ��յ�Bϸ�����������ӣ�NF-��B��������צ��������յ�NF-IL-6ת¼���ӻIL-6�����ӡ�
������6���̼�����ϸ���;���ϸ������IL-6��TNF����ͨ������ϸ���;���ϸ������IL-8�鵼��������ϸ�����������á����⣬IL-1�յ���Ƥϸ������̼�������ϸ���ͷ���֢������֢���ʣ�ֱ�Ӳ�����֢�������̡�
����IL-1��TNF����ѧ���ʣ������ڷ����������÷����Ϊ���ƣ�����4-8������TNF-���кͿ����Ԥ���ڶ���������ݿˣ�ͬʱ������IL-1��IL-6ˮƽ��������ijЩ�����£�IL-1ˮƽ�ܵ�TNF�ĵ��ء�
����6.IL-1���ٴ�
������1�����ȣ�IL-1��һ����Դ����ԭ�ʣ�endogenous pyrogen), IL-1�������������ڶ��ز�ͬ��IL-1�������У�����ע�䲻�������ܡ�IL-1�����ȵĻ���֮һ������IL-1�ٽ�����-����ϸ���ͷ�PGE2���̼����������µ������࣬�����ȡ���˹ƥ�֡�����ʹ������PGE2�ĺϳɣ�����Ϊ���ȼ���PGE2��IL-1�����и��������á������˻�֢����PBMC�е���ϸ������IL-1�������������ˣ��������Ⱦ���׳������Է��ȵ��ٴ�֢״�йء�
������2���ٽ���ϸ���ϳɼ����ڵ��ף�acute phase protein)����C-��Ӧ���ס�Ѫ�������A���ף�serum amyloid a protein,SAA)�ͦ������ǵ��ף���-acid glycoprotein����AGP���Լ�ijЩ�������֣������ڻ���ֿ���ԭ����ȣ��ǻ���������Է�������֮һ���ͼ���IL-1����߶�������ͺͶ���ű���ĵֿ�����
������3���Լ���ϸ��������ϸ�������ã����ʪ�ؽ��ؽ����ھ���ϸ���ܵ��̼��ͻ��ɷ���IL-1���̼���Ĥϸ��������ϸ��������άϸ�����ڴ���PGE2����ԭø�����Ե���ø�ȣ��Ӷ�ʹ�ؽ��еĽ�ԭ��֯���⣬�������գ��ֲ�Ѫ��ͨ�����ӣ�ֱ�Ӳ���ؽڵIJ������ˡ����ֹؽ��Ĺؽ�Һ�пɲ����ˮƽIL-1��IL-1���ɴ̼�����֯����ϸ���������γ��ۣ������������йء����⣬IL-1�̼���С��ϵĤϸ��������������ʵ����ά�������������Խߡ�
������4�����������ã�����IL-1��ЭͬIL-2��IFN-���յ�CTL��NKϸ����ɱ�˻��ԣ��ٽ�ɱ��ϸ��IL-2����ı����Լ�����άϸ������IL-6����˾��п��������á��ٴ�������IL-1����������
������5�����������ã�radioprotective)��С��ע��IL-1100ng��950R���������¿���߰�ϸ��ˮƽ���ӳ����ʱ�䣬��Ԥ��������ɵĹ������ơ��ٴ���������IL-1���ƹ�����ֲ������ڶ���ʵ���з���IL-1�ܽ���Ѫ��ˮƽ���п��ܳ�Ϊ����������ҩ��
����������IL-2
����1976��Morgan�ȷ���С��Ƣϸ�����������к���һ�ִ̼�����ϸ�����������ӣ��������������ܴٽ���ά��Tϸ��������������ΪTϸ���������ӣ�t cell growth factor,TCGF����1979��ͳһ����Ϊ��ϸ������2��interleukin 2,IL-2)��
����1.IL-2�IJ�����IL-2��Ҫ��Tϸ����Tϸ��ϵ������
������1��CD4���Ի�CD8����Tϸ������˿����ԭ�̼�CD4���Ի�CD8����Tϸ����Ⱥ���ɲ���IL-2��ͬ�����忹ԭ��Ҫ�̼�CD4����Tϸ������IL-2��PBMC��Ƣ�ࡢ�ܰͽ�ͱ������е�Tϸ���ܵ��̼����ܲ���IL-2����С��Thϸ���У�ֻ��Th1��Ⱥ�ɲ���IL-2��
������2��Tϸ������ϸ��ϵ���Ѫ��ϸ��ϵ���˺Ͷ���ijЩTϸ����Ѫ��ϸ��ϵ������ϸ������˿����ԭ�����������壨��A23187����PMA�̼��¿ɲ�����ˮƽ��IL-2���糤��ԳTϸ��ϵMLA144���Է�����IL-2��С��������ϸ��ϵEL-4����Jurkatϸ����ֹ״̬���ϳɺͷ���IL-2���̼���ɷ��ڸ�ˮƽ��IL-2��
������3��T�ܰ�ϸ���ӽ�����T�ܰ�ϸ���ӽ���123��FS6-14.13��HT-24A����ConA�̼��²���IL-2��
������4��Ӧ�û��̼����Ʊ���1983��Taniguchi�ȴ�ConA�̼���Jurkat��Ѫ��Tϸ���п�¡�ɹ�IL-2cDNA�����ڴ˾��еõ���ˮƽ�ı��ĿǰӦ�û��̼������Ʊ��ʹ�����IL-2�������ٴ�����ijЩ����������������
����2.IL-2�ķ��ӽṹ�ͻ�����IL-2����133������л���������Ϊ15.5kDa����ȻIL-2��N�˺����ǻ������ǻ���IL-2������ѧ����������Ӱ�죬�ȵ����6.6��8.2��IL-2���Ӻ���3�����װ��ᣬ�ֱ�λ�ڵ�58��105��125λ�����ᣬ����58λ��105λ���װ���֮�����γɵ����ڶ�������ڱ���IL-2����ѧ��������Ҫ���á���IL-2���������ᴿ���Թ����У��������������Ӽ��γɶ�������ή��IL-2�Ļ��ԡ�������Ӧ�õ�ͻ�䣬����125��λ���װ���ͻ��Ϊ�������˿������ʹֻ���γ�һ�ֶ��������֤����IL-2���Թ��̵Ļ��ԡ����б����õ����̼�����������rIL-2����IL-2���ӵ�125λ���װ����Ϊ�����ᣬ�Ĺ���IL-2�Ȼ��Ա���ȻIL-2�������ӡ���IL-2����λ�ڵ�4��Ⱦɫ�壬��Լ5kb����4�������Ӻ�3���ں�����ɡ��˺�С��IL-2����DNA������63%ͬԴ�ԡ�
����3.IL-2�����塡IL-2R���ɦ����ºͦ���������ɡ�
������1��IL-2R������Uchiyama(1981)�״��Ʊ��˿��Tϸ����ԭTac��McAb����IL-2�������ϵ�Tac����ϸ����Tac�ķ�����Ϊ55kDa��1984��Leonard��Tac���ӵ�cDNA��¡�ɹ���Tac����Ϊ�ǵ��ף���272�������л���ɣ�����21��������л��ź��ģ�������Ӻ�251�������ᣬ���ж�����װ��ᣬ2��N-�ǻ���λ�㣬��Ĥ���Ͱ������ֱ�19��13��������л�����Tac�Ļ���λ�ڵ�10��Ⱦɫ�壬����8�������Ӻ�7���ں��ӣ���Լ25kb��Tac��p55����ΪIL-2������������ǵ�λ�����ֳ�CD25���ǻT�ܰ�ϸ���ı�־���ڹ�����ֲ�����ȥTac���Թ���ϸ�����Խ�����ֲ�������Ӧ��GVHR�������ѽ�������ٴ���֤��Ҳ���ÿ�IL-2r McAbѡ���Եط�ա��������ЧӦϸ�����Ӷ�����ͬ��������ֲ���ųⷴӦ��ijЩ���������Լ�����
������2��IL-2R������������70kDa�����ֳ�p70�����˰�ϸ���ֻ���ԭ�б��ΪCD122����IL-2R��������λ��22��Ⱦɫ�塣����IL-2R������525�������ᣬ5��N�ǻ���λ�㣬������Ĥ��������Ĥ���Ͱ���������Ĥ������214����������ɣ���8��Cys����ṹ����1����ϸ�������أ�EPO�����峬���������ԵĽṹ����1��������άճ�����ṹ��Ĥ��25�������ᡣ��������286�������ᣬ��EPO�����������һ����ͬԴ�ԡ�IL-2R�����������Ұ��ἤø���������������������ṹ��һ���ǿ���Ĥ�˵�˿���ḻ��������IL-2�յ�����ֳ�źŴ���������Ҫ���ã���һ�������Ұ��ἤø��������������ȱ�����������IL-2R����ͻ�����ܴ�����ֳ�źţ����յ�ת¼c-myc,���ܽ鵼�յ�ת¼����Fos�����ã�ȱ��˿���ḻ������IL-2R����ͻ���岻���յ�ϸ����ֳ��c-myc��ת¼����ˣ��Ұ��ἤø;���ƺ���c-fos ������յ��йأ����Ǽ�ø������;����c-myc������յ��йء�IL-2R������Ҫ�ֲ���Tϸ����������ܰ�ϸ����LGL����Bϸ����pre-Tϸ����
������3��IL-2R�ã��ǵ��ף���347�������ᣬ������64kDa����Ĥ�ṹ�������ں�ϸ�������ؼ����Ա����������86�������ᣬ��288��321λ�����������ƺ�ͬԴ��srcͬԴ��2��SH2)����������һЩ���ữ���������ữ�Ұ���л������������źŵ�ת����IL-2R���������ڶ����ܰ���ϸ�����棬��Molt-�¡�Molt-4��Jurkat��MT-1��MT-2�Լ�EB������Ⱦ��Rajiϸ����
������4��IL-2R������������Ĺ�ϵ������IL-2R�������ܽ��IL-2��������������IL-2R���¦�������������IL-2R�����¦���������ɡ�IL-2���ڻ��Լ��ź�ת���DZ���ġ�X-������֢��������ȱ��֢���˵�IL-2R�û�����ͻ���ɥʧIL-2R���ܡ�
��4-2 ��������IL-2R�����
| ��� | ������Kd) | ϸ���ֲ����� |
| ������p55,CD25) | �ͣ�10-8M | B�ܰ�ϸ�� |
| ������p70,CD122)+���� | �У�10-9M | YT��NKϸ���꣩��MLA144 |
| ����+����+���� | �ߣ�10-11M | PHA�̼�ĸϸ����HUT102B2 |
������5��������IL-2R��������IL-2R��soluble IL��2��receptor,sIL-2R����Ĥ�����ʽIL-2R�����������������45kDa��������Tϸ����Ѫ�����Ͳ�����HTLV-I����Ⱦ��HUT102B2ϸ�����������к��д���sIL-2R��PBMC��˿��ԭ��CD3McAb��ͬ�����忹ԭ�̼�����ͷ�sIL-2R��������Ѫ�����Һ����ɼ������sIL-2R��sIL-2R������Ĥ����IL-2R��mIL-2R���������IL-2���Ӷ���Ϊһ�������������ʡ�sIL-2R���߿ɼ���ijЩ�����������������߲���������Ⱦ�Լ����Լ���ֲ�ų�ȣ������µ����ڣ���
����4.IL-2������ѧ���á�IL-2�����þ�������ϵ��������Լ���ԣ�������Լ���Ե��ص㣬���˵�IL-2�ܴٽ�С��Tϸ������ֳ����С���IL-2����ά����Tϸ����������IL-2���ڵİ�˥��ֻ��6.9���ӡ��б�����PEG��IL-2�������Σ�������ѧ������Ӱ�죬��˥�ڿ��ӳ�7�����ҡ�Ŀǰ����IL-2������ѧ���ô�������ʵ��Ľ���������кͻ��ԵĿ�IL-2���������IL-2������ѧ���ԡ�
������1��Th��Tc��Tsϸ������IL-2���ķ�Ӧϸ����IL-2�Ծ�ֹTϸ�����ý���������ϸ����Tϸ������ԭ����˿����ԭ��ͬ�����忹ԭ�̼��������IL-2���ڵ������½���S�ڣ�ά��ϸ������ֳ��IL-2�ɴ̼�Tϸ��ת���������壨TfR��CD71�����ȵ������塢MHC���ԭ�ı�������������ܰ�������IFN-�á�IL-4��IL-5��IL-6��TNF-�¼�CSF�ȡ�
������2���յ�CTL��NK��LAK�ȶ���ɱ��ϸ���ķֻ���ЧӦ���ܣ����յ�ɱ��ϸ������IFN-�á�TNF-����ϸ�����ӡ�IL-2����ǿCTLϸ�������أ�perforin)����ı��
������3��ֱ��������Bϸ�����ٽ�����ֳ���ֻ���Ig���ڡ��ѷ��ֻ��Bϸ��Ҳ�ɾ���IL-2R��IL-2��Bϸ���ĵ������ó�ͨ���̼�Tϸ������Bϸ����ֳ�ͷֻ������⣬��������ֱ�ӵĵ������á�
������4�������ϸ����
����5.IL-2���ٴ�Ӧ�á�Ŀǰ����IL-2�������ٴ����������Լ���Ⱦ�Լ����ȡ�
������1����������IL-2��������յ�PBMC�����������ܰ�ϸ����TIL����Ϊ�ܰ����Ӽ����ɱ��ϸ����LAK����LAK/IL-2����ϸ���������������Ǻνܽ����ܰ������᳦ֱ�����н�������Ч���Ըΰ����ѳ�����ͷ�����۰������װ����ΰ����в�ͬ�̶ȵ���Ч���������ѿ�ʼ����IL-2�������ƶԺ���������ϸ�����Լ���ĸϸ�����������Ƚ����ٴ���֤��
������2�����Ƹ�Ⱦ�Լ���������ʵ����������IL-2��ijЩ��ϸ�����߹��ܵ��¶��ܲ�����Ⱦ������ǿϸ�����߹��ܵIJ�����һ����Ч��IL-2������ֱ�ӿ��������ԣ�����ͨ����ǿCTL��NK�����Լ��յ�IFN-�ò������鵼��������Ⱦ�ġ�Ŀǰ��IL-2���ƻ�Ը�������ʾ����ϲ����ͷ�����ڵ����������Ⱦ��AIDS�����ѽ�������ٴ���֤�����������硢��˸˾���Ⱦ��Ҳ��һ����Ч����rIL-2�����ӳ���˸˾�H37RV���ȾС�������İ�������ʱ�䣬���������ʣ����ٸ�Ⱦ����Ƣ������֯�ڵĽ�˸˾�����
������3�������������ã�adjuvanticity):Ӧ��IL-2��Ϊ����������ԭ�������ǵ�λ��������Ӧ�ã�������屣��������Ӧ���ˮƽ�����⣬�������IL-2���н���Ѫѹ���ã�IL-2���Ƹ�Ѫѹ�ѽ�������ٴ���֤��
������4������IL-2�������ںϵ����������ʪ�Թؽ������/�����ٴ���֤��Լ��74%���߲���õ����ƣ�IL-2�ں϶�����Ҫ������CD4�����ܰ�ϸ�����нϺõ�ѡ���ԡ�
����rIL-2���ڴ����ʹ�ö������ýϴ�����ëϸѪ����©�ۺ�����capillary leak syndrome��CLS�������⣬IL-2��˥�ڶ̣���ʱ�����������յ�����һ�����塣
��������ImmunexӦ�ÿ�IL-2R����McAbԤ��������ֲ��GVHR��������ٴ����顣
����������IL-3
���������ڼ���̼����ӡ�
�������ģ�IL-4
����1982��Howard����Tϸ��������������һ�ִٽ�Bϸ����ֳ�����ӣ��������ΪBϸ����������-1��b cell growth factor-1,BCGF-1)���е�ʵ���ҳ�ΪBϸ���̼�����-1��b cell stimulating factor-1,BSF-1)��Tϸ����������-2��T cell growth factor-2,TCGF-2����1986������¡�ɹ�������ͳһ����Ϊ��ϸ������4��interleukin 4,IL-4)��
����1.IL-4�IJ�����������IL-4��Ҫ�ɻTϸ����������С����Th2��Ⱥ���������⣬�ʴ�ϸ����IL-2�̼�С��Tϸ��ϵ2.19��ConA�̼���Th��¡2F1��С��������EL-4ϸ���Լ�Bϸ��ϵCH12���ܷ���IL-4��
����2.IL-4�ķ��ӽṹ�ͻ���С��IL-4����Լ6kb������IL-4������120������л���ɣ����ķ�����Ϊ14kDa����3���ǻ����㣬���ǻ�����IL-4������Ϊ30kDa����IL-4����λ�ڵ�5��Ⱦɫ�壬��4�������Ӻ�3���ں�����ɣ�Լ10kb������֪�ܰ����ӻ����нϴ��һ����������IL-4������129������л���ɣ�15kDa����2���ǻ����㣬����6�����װ��ᣬ��������ڶ��������ɡ�������IL-4DNAˮƽ����70%ͬԴ�ԣ�IL-4ǰ�嵰�״�N�˵�91λ�������Լ�C�˵�128λ��������С��֮���ڰ�����ˮƽ����70%ͬԴ�ԣ�ǰ�嵰��91��128λ������֮�������ͬԴ�ԣ��������IL-4���������������йأ����ˡ���IL-4����ѧ������û�н��淴Ӧ��
����3.IL-4�������ˡ�IL-4���壨IL-4R����800������л���ɣ�������Ϊ140kDa����Ĥ����207�����ᣬ��Ĥ��24�����ᣬ������569�����ᣬ��С��IL-4R��53%ͬԴ�ԣ����ں�ϸ�����������峬�����Ա���������ΪCDw124����С��Tϸ����Bϸ��������ϸ��������ϸ��������ϸ���ͷʴ�ϸ�����涼��IL-4R��ÿ��ϸ��������Ŀ��100��2000���ң�������Kd��10-10��10-11M��1��5��10-12MŨ�ȵ�IL-4��ʹBϸ����ֳ��3H-TdR�����ʣ��ﵽ���ֵ��50%��LPS��Bϸ��IL-4R���������������ã���������������5��10����IL-4����Ӧ�����Ϻ�ϸ���ڴ����źŵ�;���в������IL-4R�����������հ����˿���ᣬPTK��PKC���ܲ�������鵼���źŴ��ݣ�IP3��Ca2+Ũ�����ߡ���С�����в�ͬ������ʽmRNA������ķ����ͣ������ԣ�IL-4R(sIL-4R����sIL-4R��Ĥ��IL-4R��mIL-4R��ͬ����IL-4���ʱ������ͬ��������sIL-4R������IL-4���������壬�����IL-4������ѧ���ԡ�
����4.IL-4������ѧ���ԡ�IL-4����Bϸ����Tϸ�����ʴ�ϸ��������ϸ������ϸ���������ߵ������á�
������1��Bϸ�����ٽ�SAC��IgMԤ�ȴ̼�Bϸ������ֳ����һ����ѧ�����ѱ�������Ϊ���IL-4������ѧ���ԡ�IL-4�ٽ�Bϸ��MHC���ԭ��Fc��R��/CD23��CD40�ı������ǿBϸ����ʿ�ԭ������ʹ����ϵͳ��С����ԭ�̼���������Ӧ������Fc��R��/CD23��Ige Fc�ε��������壩�ı�����ͷſ�����CD23��sCD23��/IgE������ӣ�IgE-BF������mIgE����ϸ����ϲ��յ���ֻ���������ٽ�Bϸ��IgE�IJ����йء�IL-4���LPS�̼�С��Bϸ��IgG1��IgE����ˮƽ�ֱ�Ϊ8��10����10��100��������IgG3���ڽ���6��10����IgG2a��IgG2b��IgM�в�ͬ�̶��½�����IgA����������Ӱ�졣IL-4��ǿIgG1��IgEˮƽ�Ļ��������ǣ���ʹѡ���Դ̼��������IgG1��IgE��Bϸ������ֳ�ͷֻ�������������������ȶ�����CH��Ig���ת����S��-S��1��ϣ��ٽ�IgG1�ϳɺͷ��ڡ�IFN-�ö�IL-4��������ѧ�����������������á���IL-4�������IFN-��mRNA��ת¼������IFN-���յ�Bϸ������IgG2a���������ȾʱѪ��IgG1��IgEˮƽ���ߡ����⣬IL-4���ɴٽ���ֹ��Bϸ�������ڻ����Go�ڽ���G1�ڣ�ϸ�������������CD25��
������2��Tϸ����IL-4��Tϸ���������ڵ��������ӣ���HT-2ϸ��ϵ��һ��IL-2����ϸ��ϵ��IL-4�ɵ���ά��TH-2����ֳ����IL-2�Ϳ�IL-4McAb��11B11���ɷֱ�����IL-2��IL-4�̼�IL-2ϸ������ֳ���ã����֮�������������á��������Tϸ����CD4���Ի�CD8������Ⱥ����IL-4����ֳ��Ӧ����ҪIL-1����PMA����Эͬ���ã���IL-2������ά�ֻTϸ������ֳ������IL-4�����źŵĴ��ݹ�����IL-2������ͬ������IL-4���յ�CD4-CD8+��CD4+CD8-��CD4-CD8-����ϸ������ֳ��IL-4��ǿPHA�̼�Tϸ���ͷ�GM-CSF��G-CSF����С��ʵ��ϵͳ�У����������IL-4��CTL��LAKϸ���ķֻ������������ã������˵�ɱ��ϸ������ʱ����Ϊ���������ã�������MLC��IL-4ѡ���Եشٽ�Th��ֳ��ͬʱ����CTL��NK��LAK���ܵĽ��͡�����IL-4��������IL-2���յ���NK��Ѫ��ϸ��LAK���ԡ�������֣���IL-4��Tϸ����Bϸ���������ɽ�������IL-2Rλ����������Ӱ�������IL-2R��������IL-4���ɴ̼�CD3����NKϸ����¡��������
������3���̼��ʴ�ϸ����ֳ������IL-3��Эͬ���ã��������ճĤ�ͽ����֯�ͷʴ�ϸ�����������DZ���ġ�
������4���ٽ�����ϸ����ʿ�ԭ��ɱ������ϸ���Ĺ��ܣ����������MHC���ԭ��FcR�����йء�IL-4��GM-CSF��IL-3��LPS��Эͬ���á�IL-4���յ�����Ѫ����ϸ������G-CSF��M-CSF����ǿ������ϸ���鵼�����ɡ�ɱ�˻��Ժ�ADCC���á�IL-4��С�����ϸ���������ӣ����ٽ�IL-1ra�����������Ƶ���ϸ��IL-1��TNF��IL-6�IJ�����
������5��ЭͬCSF�̼���Ѫϸ������ֳ����G-CSFЭͬ��ǿ��ϸ�������γɣ�Эͬ��ϸ�������أ�EPO����ǿBFU-E���γɡ�
����IL-4��Ϊ�������ߵ��ڼ��ѽ�������ٴ����顣���⣬����ʼ������������ȱ��֢���ٴ����顣�������ڡ������֤ʵIL-4��������IL-1��IL-6��TNF���ڣ����ٽ�IL-1ra ���������Ӧ��IL-4����Ϊ���ư�Ѫ֢�ݿ��ṩһ���µķ�����
����(�壩IL-5
����1980��Takatsu�ȷ�����Tϸ����������Һ�У�����һ�����������Tϸ��������Эͬ����������ԭ�Ŀ���Ӧ�𣬳�ΪTϸ��������ӣ�t cell replacing factor,TRF)�������������Ӷ�Bϸ���͆�������ϸ����ֳ���ֻ�����Ҫ�������ã�����Bϸ������������-��b cell growth factor-��,BCGF-��),IgA��ǿ���ӣ�IgA-enhancing factor,IgA-EF������������ϸ������̼����ӣ�eosinophil colony-stimulating factor,Eo-CSF������������ϸ���ֻ����ӣ�eosinophil differentiation factor,EDF)��1986��ͳһ����Ϊ��ϸ������-5��interleukin 5,IL-5)��
����1.IL-5�IJ�����������IL-5��Ҫ�ɻTϸ����������С������Th2��Ⱥϸ��������
����2.IL-5�ķ��ӽṹ�ͻ���1986��Kinashi���IL-5cDNA��¡��С��IL-5��133������л���ɣ���21��������ź��ģ�����IL-5���Ӻ���112������л������ķ�����12��15kDa����3���ǻ���λ�㣬�ǻ����������Ϊ18kDa���ǻ�������IL-5���Ա����Լ�����Ӧ����Ľ������Ҫ���á�С��IL-5ͨ���Զ�������ӵĶ�������ʽ���ڣ�������Ϊ45kDa���˵�IL-5��134������л���ɣ���22������л��ź��ģ�2���ǻ����㣬�˺�С��IL-5����ֱ�λ�ڵ�5�ź͵�11��Ⱦɫ�壬��IL-3��IL-4��GM-CSF����Ѫ���ӵĻ��������������˺���IL-5�ڰ�����ˮƽ����70%��ͬԴ�ԣ�����ѧ�����н��淴Ӧ��
����3.IL-5���塡С��IL-5�ɦ��ͦ���������ɡ�������p60����415�������ᣬ�ǵ��ף��ȵ�����17�������ᣬ��Ĥ����322������л�����Ĥ��22������������54��������л��������������IL-5Ϊ�������������źŵ�ת����������p130�����������IL-5���������ͬ��ɸ��������塣С��IL-5R����IL-3R�Ħ�����AIC2B��������ͬ����IL-5R��������IL-3R������GM-CSFR���ǹ�ͬ�ġ�����mRNA���ӵIJ�ͬ���ѷ����ж��ֿ�����С��IL-5R����������һ�ֿ�����IL-5��Ĥ���IL-5R�Ľ�ϡ��˺���IL-5R������79%��ͬԴ�ԡ�
����4.IL-5������ѧ���ԡ�������IL��ȣ�IL-5����ѧ������������Խ�խ��
������1��С��IL-5�ٽ���ԭ�̼���Bϸ���ֻ�Ϊ����ϳ�ϸ������Ҫ�����ڽ���ϸ����ֳ���ڵ�Bϸ���������ӻBϸ��IL-2R�ı��IL-5�����ִ̼���������IL-6�������ƣ���IL-5ֻ������Bϸ���̼����խ��ʱ���ڡ�
������2���ٽ�IgA�ϳɣ�����������ǣ�����ΪIgA�������������ӣ�ʹmIgM ����Bϸ���ֻ�ΪmIgA����Bϸ������������IgA��Bϸ�����ٽ�����ֳ�ͷֻ�����Ϊ����IgA�Ľ�ϸ����IL-4��ЭͬIL-5�ٽ�IgA�ϳɵ����á�IL-5��IgM�ķ���Ҳ�дٽ����á�
������3��ЭͬConA��IL-2�յ�������ɱ����Tϸ��ǰ�壨CTPp���ֻ�ΪCTL��
������4����������������ϸ�����ӳ�������������ϸ���Ĵ��ʱ�䣬�̼��˺�С����������ϸ���Ĺ��ܣ��յ���������ϸ���ķֻ���
����������IL-6
����1980�귢�ֳ���άϸ����Poly I-C�̼����ܲ���һ�����Ʋ������Ƶ�ϸ�����ӣ���Ϊ��2�����أ�IFN-��2�����Ժ���о����δ��֤ʵ�������ӵ�ֱ�ӿ��������ã�������������������ѧ���ܣ�����ʵ��ϵͳ���ܵIJ�ͬ����������Ϊ�ӽ���/��ϸ�����������ӣ�hybri-doma/plasmacytoma growth factor,HPGF),Bϸ���ֻ����ӣ�B cell differentiation factor,BCDF),Bϸ���̼�����-2��b cell stimulatory factor 2,BSF-2),26kDa,��ϸ����Tϸ���ֻ����ӣ�cytolytic T cell differentiation factor,CDF)��ϸ���̼����ӣ�hepatocyte stimu-lating factor,HSF)�ȡ�1986��ͳһ������ϸ������6��interleukin 6,IL-6)��
����1.IL-6�IJ������ܰ�����ijЩ���ܰ���ϸ�����ɲ���IL-6����
������1��Tϸ����Tϸ������IL-6�����ھ���ϸ����PMA����ԭ���ϸ���̼���Ӧ��Tϸ����¡���Լ�HTLV-I��Ⱦ��Tϸ��ϵ�Ⱦ��ɷ���IL-6��
������2��Bϸ������SAC�̼������Bϸ����
������3������ϸ����LPS�̼�����ϸ������IL-6��ijЩ����ϸ��ϵ��P388D1Ҳ�ɷ���IL-6��
������4������άϸ�������Է�����IL-6���������ӻ�̼�����IL-1��TNF��PDGF��IFN-�¡�PolyI-C��A23187��PMA�ȿɴٽ�IL-6�IJ�����
������5����С��ϵĤϸ��������ϸ������Ƥϸ������һ�����������¾��ɲ���IL-6�����⣬����ϸ����ϸ��ϵ��MG63�ɹ�������T24���װ���A549�ΰ���7860������SK-MG-4����ĸϸ������U373��״ϸ����������ճҺ��ϸ��������ϸ����Ҳ�ܷ���IL-6��������ִ���ǰҶ�е����ݡ���״ϸ����folliculostellate)�ɲ���IL-6���������Ѫ֢ʱLPS�̼�����GH��ACTH�ȼ���ˮƽ�����йء�
����IL-1��TNF��IFN-�¡�PDGF��LPS��Poly ��-C��A23187��PMA�ȶ�IL-6�IJ����������������á�
����2.IL-6�ķ��ӽṹ�ͻ���1985��Kishimoto�ȴ���Tϸ�������Ȼ��IL-6cDNA��¡�ɹ�����IL-6������С����65%ͬԴ�ԡ���IL-6����λ�ڵ�7��Ⱦɫ�壬��Լ5kb,��5�������Ӻ�4���ں��ӡ�
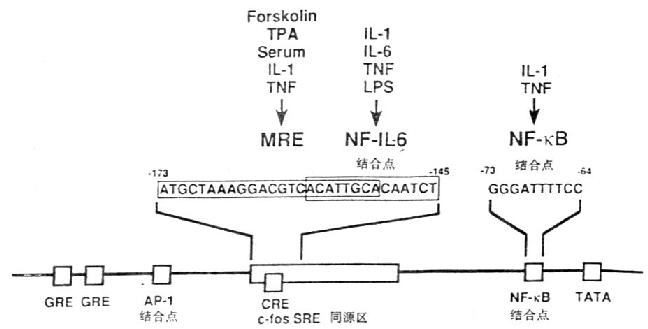
ͼ4-1��IL-6����Ĺ��ܵ�����
������IL-6�����ܵ����������д����Ŷ���ת¼����Ԫ����transcriptional control element)������Ƥ�ʼ��ط�ӦԪ����glucocorticoid responsive elements,GRE)��AP-1���λ�㡢c-fosѪ�巴ӦԪ��ͬԴ�c-fos serum responsive element homology,c-fos SRE homology)��cAMP��ӦԪ����cycli AMp responsive element,CRE)��NF-��B���λ�㡣IL-1��TNF��ϸ�����ӿ�ʹIL-6�����Ӻܿ췢��һ���ԵĻ��IL-1��Ӧ��Ԫ����IL-6��������-180/-123��IL-6�����ӣ�NF-IL-6��ʶ��һ�������14bp,ACATTGCACAATCT���෴ӦԪ����multi-response element,MRE��λ��c-fos SREͬԴ���ڣ���������IL-1��TNF��forskolin��PMA�յ�IL-6�����йأ���IL-1��TNF�̼�IL-6�����йص�NF-��Bλ��TATA�е����Ρ�
������IL-6������212��������л���ɣ�����28��������л����ź����У�����IL-6Ϊ184������л���������26kDa��IL-6������4����������C�ˣ�175��181λ�����ᣩ�����ϵ�����ɣ�����179λ������л�����������Ľ�Ϸdz���Ҫ���������ǻ�������ѧ���Թ��ܲ��DZ��裬N��23��������л��䲻ֱ����IL-6����ѧ�����йأ���������IL-6����������ȶ����á���IL-6������������С��IL-6��42%ͬԴ�ԣ��˵�IL-6��С��ijЩϸ���д̼����á�IL-6��G-CSF��IFN-���нϸ�ͬԴ�ԣ��Թ�����Ѫϸ����������Ѫ��ϸ����ijЩ����Ҳ������֮����
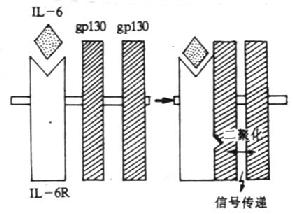
����3.IL-6�����塡Ŀǰ��֪��IL-6R�����ɳ�֮ΪIL-6������嵰�ף�IL-6binding receptor protein���ͳ�Ϊ�ź�ת�����ף�signal-transducing protein����gp130����ɣ�ϰ����ǰ�߳�֮IL-6R��
������1��IL-6R��CD126������IL-6R��468����������ɣ��г�N��19��������л���ij��������449�����ᣬ��Ĥ��������Ĥ���Ͱ������ֱ�Ϊ339��28��82�������ᣬ������Ϊ80kDa��6��N�ǻ���λ�㡣��Ĥ����һ��Ig������C2��Լ100�����ᣩ��2��������ά��ϵ��ṹ������100�����ᣩ��1��ϸ�����������ͬԴ������ɣ����ߺ�4�����ص�Cys��һ��WSXWS�ṹ������IL-6R��IL-6���Ϊ��������IL-6R�ֲ����ܰ���ϸ���ͷ��ܰ���ϸ������Bϸ����EBVת��Bϸ���������ܰ�ĸϸ����Ѫ��ϸ����������ϸ������ֹTϸ������ϸ��������ϸ��������������Ѫ����AML��ϸ�����ȸ�ϸ����ϸ���ȡ�
������2��gp130(CDw130����������Ϊ130kDa���ǵ��ף�����14��DZ��N-�ǻ���λ�㣬��Ĥ��������Ĥ���Ͱ������ֱ���597��22��277�������ᡣ��Ĥ������1��IgC2����6��������ά��ϵ��Ľṹ�����еڶ����͵������ṹ��֮����4�����ص�Cys��WSXWS�ṹ�������γ�1��ϸ�������������ṹ�����Ľṹ��gp130����ֱ�������IL-6��ϣ�����������£�IL-6��IL-6�����Ϻ�ʹIL-6R�Ĺ������仯��Ѹ��������gp130���ӽ�ϣ��γɸ������Ľ��λ�㣬��ͨ��gp130�ǵ�λ�����źš��˺�С��gp130�ڰ�����ˮƽ����77%��ͬԴ�ԡ�תȾgp130cDNAС��pro-Bϸ����IL-6/sIL-6R������̼��¿ɴ�����ֳ�źš�С������ע��IL-6������gp130mRNA�ı��Ŀǰ��֤ʵ��gp130�����IL-6�����������⣬Ҳ�ǰ�Ѫ���������ӣ�LIF����������M��OSM������״��Ӫ�����ӣ�CNTF����IL-11�����������õ��ǵ�λ��
������3���ź�ת����gp130��IL-6/IL-6R�������Ϻ̼�gp130���ڲ��ַ����Ұ������ữ��Ŀǰ���ڲ���˹��̵��Ұ��ᵰ��ø�����û���������ᰱ�ἤø�������̶�����˿����/�հ��ἤø��˿��ԭ��ĵ���ø��mitogen activated protein kinase,MAPK)�ļ��ʹNF-IL-6��˿������հ������ữ��������Ӷ��ٽ���Ӧ����Ļ��
������4��sIL-6R��������������������ϸ��ϵU266�������壬PHA���PBMC�Լ�HTLV-I����ϸ��Ҳ�ܷ���sIL-6R��������Ϊ50kDa���÷�ת¼PCR��������ϸ��������ϸ���о����������sIL-6r mRNA,���з���������Ĥ���������Ӧ��������һ�¡�sIL-6Ҳ�ɴ�Ĥ��ϵ�sIL-6R��mIL-6)���������sIL-6R��IL-12p40�ǻ����и߶�ͬԴ�ԣ���IL-6��IL-12��p35�ǻ����и߶�ͬԴ����˿����Ʋ�������IL-6/sIL-6R�������IL-12���ӿ���Ҳͨ��������pg130����������ϸ����������������ϸ���������岻ͬ��sIL-6R���IL-6�����ϸ��Ĥ����gp130��ϣ���ǿIL-6�Ĵ̼����ԡ���������gp130��sgp130��������sIL-6R/IL-6������Ļ��ԡ�sIL-6Rˮƽ��������ijЩ���������Լ����йء�
����4.IL-6������ѧ����
������1���̼�ϸ��������IL-6�ɴٽ�����ϸ������ֳ����B�ܰ�ϸ���ӽ�������ϸ������EBVת����Bϸ����Tϸ����PMA��IL-4�̼�������ϸ������Ѫ��ϸ��������ϸ������С��ϵĤϸ����
������2���ٽ�ϸ���ֻ�����Bϸ���ֻ���Ig�ķ��ڣ�CTL�ֻ���ЭͬIL-2��ǿCTL�д����ػ���ı��������Tϸ��IL-2������IL-2R����յ������ϸ������ϸ����NKϸ���ֻ���ЭͬIL-3�ٽ���ϸ���ֻ��;�ϸ���ij��졣���Դٽ�С�������ֲ�����߹��ܵ��ؽ���
ͼ4-2 IL-6�����źŴ��ݵ�ģʽͼ
������3�����ٸ�ϸ�������ڵ��ף�acute phase protein)�ĺϳɡ�
������4������M1������Ѫ��ϸ��ϵ���������ٽ������ͷֻ������ƺ����������ٰ�ϸ��������
����5.IL-6���ٴ���IL-6���ٴ��϶��ּ����ķ�����һ���Ĺ�ϵ��
������1��IL-6�����������Լ���
����������ճҺ�A��������������Ϊ�߱���Ѫ֢���ж���Ѫ�������Լ������ڵ������ߡ�������ճҺ��ϸ������IL-6��IL-6mRNA������Ѫ����IL-6�������ߣ������пɼ���IL-6�����Ķ��¡��ϸ����ֳ������������֢״����֢�ɼ������ˡ�
������Castleman�ϲ������߱���Ϊ�߱���Ѫ֢�������ڵ���ѪС�����ߣ��������ܰͽ�����������B�ܰ���ϸ������IL-6������ijЩ���߿ɷ�չ��Ϊ��Թ�������
���������ʪ�Թؽ��ף�����Ϊ���¡�Խ�ϸ������֢���������塢C��Ӧ���ף�CRP����ѪС�����ߡ�������Ѫ���Լ��ؽڵĻ�Һ���ܲIL-6����Һ��IL-6��IgG�Լ�Ѫ����IL-6��C��Ӧ����֮�������Ե�����ԡ����ߵ�Tϸ����Bϸ������Ĥϸ���Լ�����ϸ�����ɲ���IL-6��
�����ܰ��̲����밬�̲����߶��¡Bϸ����йأ�HIV��Ⱦ�յ�����ϸ������IL-6��������Ѫ����IL-6ˮƽ�����ߡ����⣬IL-6������Koposi����������Ҫ��������֮һ������IL-6���������ڿ�����Koposi������ϸ����������
������2��IL-6������
�����ٽ�ϸ�����γɣ�������֢�յ�IL-6����ϳ������뽬ϸ�����ķ����йء�����ʯ����ֲ�鸹ǻ�̼�С���յ���֢�����յ����ϸ߱����Ľ�ϸ�������ڻ���Ҳ�ɼ�����������������ȷ��������ʪ�Թؽ������뽬ϸ�����γ��йء������⣬IL-6�ɴٽ���ϸ����������ϸ����������ijЩ��ϸ����ϸ������������IL-6�Ĵ��ڡ�
�����ڿ���ͨ���Է��ڻ������Hodgkin���ܰ����������ܰ�ϸ����Ѫ���ͼ���������Ѫ���ķ����йء�Ѫ����IL-6ˮƽ���Թ������罬ϸ����Ѫ���������س̶��йء���Թ��������߲�������ϸ������IL-6R�������ӣ�����Ѫ����sIL-6Rˮƽ�������ߡ�
������3��IL-6��Ĥ��������С�����ף�mesangial proliferative glomerulonephritis,MPG):MPG�������пɲ��IL-6��������ˮƽ�뼲���ķ�չ�йء����������Ļ�����С��Ĥϸ���ɲ���IL-6����ʾMPG������IL-6���Է����йء�
����IL-6ת����С��ɳ���ijЩ���ٴ������Ƶı仯����Ѫ���и�Ũ�ȵ�IL-6��IgG1����ϸ������֢��MPG�ķ����Լ������о�ϸ���ij��졣
������4�����˺��������Ѫ��IL-6ˮƽ���ӡ�
������5����������Ĥ�ͼ���Ĥ��С��Ľ���ϸ���ɷ��ڴ���IL-6��IL-6�ֿɴٽ��Խ���ϸ������������Ӫ�����Ӻ����������ӡ�
������6��IL-6������������-����-�������ᣬ�̼�ACTH��Ƥ�ʼ��ص��ͷ��Լ���״ϸ���ϳ��ڷ��ġ�
����IL-1��TNF-����IL-6����IJ������˿�����Эͬ���á�
����Ӧ��IL-6���Ʒ��ơ���������ѪС�����֢����֢�Լ���Ϊ���������ѽ����ٴ����顣
�������ߣ�IL-7
����1982��Whitlock�����˹��賤������ϵͳ��LTBMC)��ʹ�������п��ܶ�Bϸ��ǰ�����ֳ�ͷֻ�����������о�������Bϸ��ǰ����������ڻ���ϸ�����ϸ�������ԣ��Ʋ�������ϸ�����ܷ���һ��Bϸ��ǰ��ϸ���������̼����ӡ�1988��Namen��Ӧ��LTBMC���һ��SV40����תȾ�Ĺ������ϸ���꣨bone marrow stromal cell line)IXN/A6���ܷ���һ��ǰBϸ���̼����ӣ��������Ϊ�ܰ�ϸ�������أ�lymphopoietin-1��LP-1������С��ǰBϸ���������ӣ�murine pre-B cell growth factor)��1988��ͳһ����ΪIL-7��
����1.IL-7�IJ������ɹ������ϸ�������ٻ���ϸ����������IL-7 cDNAΪ̽�룬��С�����١�Ƣ�������ε�ϸ���о������IL-7mRNA���ڣ�����С��һ��
����2.IL-7�ķ��ӽṹ�ͻ�����ȻIL-7������ԼΪ25kDa����69��90λ������л���N�ǻ����㣬��������6�����װ��ᣬ���ܲ������ڶ�������γɣ���IL-7������ѧ��������Ҫ���á�1988��Namen����IXN/A6ϸ���꽨��cDNA�Ŀ⣬ɸѡ��IL-7��cDNA������¡��COS-7ϸ���б�����л���IL-7��С��IL-7ǰ����154������л���25������л����ź��ģ������IL-7��129������л���ɣ��������ķ�����Ϊ14.9kDa��С����ȻIL-7�����������ơ�1989��Goodwin��С��IL-7cDNAΪ̽�룬���˸ΰ�ϸ��ϵcDNA���У������С��IL-7 cDNA�߶�ͬԴ����IL-7cDNA��¡������COSϸ���еõ������IL-7����λ�ڵ�8��Ⱦɫ�塣rHuIL-7������177�������ᣬ����25���������ȵ����У�����IL-7������152�������ᣬ���ķ�����17.4kDa����С��IL-7��60%ͬԴ�ԡ�IL-7��pH�ı䣨pH2.1��8)��SDS���ȵ��������ؾ���һ���ĵֿ���
����3.IL-7���塡IL-7������С��IXN/2b����ϸ��������ڸ�������Kd��1��10-10M���͵�������Kd��4��10-8M)�����������塣ÿ��ϸ��Լ��2000��2500�����壬����15��20%Ϊ�������͡���Pre-B������ϸ��������Tϸ�����Ϻ�ijЩ����ϸ������ϸ��������IL-7R������Bϸ������IL-7R���������Bϸ��ǰ���������IL-7R���Ұ��ἤø�ź�ת��;��������أ��Ұ������ữ��IL-7R�鵼�Ŀ�Ĥ�źŲ����ʹ��ݹ����бز����ٵIJ��衣���ѷ���һ������mRNA�IJ�ͬ�����������Ŀ�����IL-7R������Ч�ؽ��IL-7��������IL-7��һ�����������о�������IL-7R���ں�ϸ���������ܱ��������Ա������1��������άճ�����ṹ��
����4.IL-7������ѧ���ԡ���IL-7��������С���ǰBϸ����С��IL-7����ǰBϸ�����̼����á�Ŀǰ��֪IL-7����Ҫ����ѧ���������¼����档
������1��Bϸ�����̼�Pre��Bϸ����B220+����������TGF-�¶Դ����������ã���������IL-4����IL-6McAb�Լ���IL-2R��IL-3R��IL-5R��McAbs�����ơ��Թ������ܰ�ϸ���ķֻ�����Ҳ�д̼����á�����IL-7��10-13M���ɴ̼�IXN/2bϸ����ֳ��
������2������ϸ�����ٽ�������CD4-CD8-��CD4+CD8+ϸ����Ⱥ����ֳ��
������3��Tϸ�����ٽ�����ܰ�ϸ��������MLC����PMA�̼�Tϸ������ֳ��ЭͬConA�̼�Tϸ������IL-2�Լ�IL-2R��ICAM-1�ı��IL-7�յ�Tϸ����ֳ��ͨ��IL-2�����Ͳ���������;����
������4���յ���PBMC����LAK���ԣ�����IL-2��Ѫ���LAK�����������仯������IL-7�����յ�LAK�������ò�����IL-2��IL-7�յ�LAK��ǰ��ϸ����Ҫ��ѪPBMC�е�NK��IL-4�������յ�����������ЧӦ����IL-4��Ѫ���ʹIL-7�յ�LAK�������4��10����
�������ˣ�IL-8
�����������������ӡ�
�������ţ�IL-9
����1988��Uyttenhove�ȱ�����һ������С��Tϸ����ϸ�����ӣ���֧��ijЩThϸ����¡����������������30��40kDa���ֳ�Tϸ����������-��t cell growth factor ��,TCGF-��)��P40����IL-9����Ǵ�HTLV-���Ⱦ��Tϸ��ϵ���������з��ֵģ��ܴ̼��˾�ϸ����Ѫ��ϸ����Mo7e����ֳ��1990������ΪIL-9��
����1 .IL-9�IJ�����IL-9�ɻTϸ������Ҫ��CD4+Tϸ����������PHA��CD3McAb��Ca2+������յ�Tϸ������IL-9��PMA��Эͬ���á�PMA��Ca2+����̼���PBMC��ת¼����IL-9mRNA�����⣬HTLV-Iת����Tϸ����C5MJ2ϸ�����ʴ�ϸ��Ҳ�ɲ���IL-9��Ŀǰ��û�м���ֹTϸ����Bϸ������IL-9mRNA�ı��
����2.IL-9�ķ��ӽṹ�ͻ����˺�С��IL-9����ṹ���ƣ���DNAˮƽ����67%ͬԴ�ԡ���IL-9������5�������Ӻ�4���ں�����ɣ�����Լ4kb����IL-3��IL-4��IL-5��M-CSF��GM-CSF��c-fms��PDGFR����λ�ڵ�5��Ⱦɫ���У���С��λ��13��Ⱦɫ�塣IL-9����5��Ƿ�������UTR������TATA���Լ�����ף�activator protein,AP)1��2��3��NF-��B,specificity protein-1��SP-1��λ�����Ƥ�ʼ��ط�ӦԪ����GRE����ʶ��λ�㡣С��IL-9���ӵ�ǰ����144��������л���ɣ���18��������л����ź��ġ��˳���IL-9��126��������л���ɣ�������Ϊ14.2kDa���˺�С��IL-9�ڰ�����ˮƽ��56%ͬԴ�ԡ�������10�����صİ��װ��ᣬ����������¿����γɸ��ӵĶ������IL-9Ϊ���Ե��ף��ж��N�ǻ����㡣
����3.IL-9���塡IL-9R�ṹ�����ں�ϸ�����������峬���壨ERS������������Ϊ��������С��IL-9R��468�������ᣬ��IL-9RΪ553�������ᣬ������53%ͬԴ�ԡ���С���ѷ�����ȱ����Ĥ���Ͱ������Ŀ�����IL-9���壨sIL-9R)��
����4.IL-9������ѧ���ܡ���������ϸ�����Ӳ�ͬ��С��IL-9���������˵�ϸ�������˵�IL-9ȴ��С��ϸ���̼����ԡ�
������1��ά��Tϸ��������Ϊһ���Է����������ӣ�����ֳ�ΪTϸ����������-��TCGF-�����ڿ�CD3McAb���յ�IL-9���������Ʋ�IL-9��Tϸ����ԭ�̼�������������Ӧ������Ҫ�������á�
������2��IL-9����IL-4�յ�PBL������Bϸ��IgG��IgE��IgM�IJ����дٽ����á�
������3��С��IL-9ЭͬIL-3��IL-4�̼�������Դ�ʴ�ϸ������ֳ�����յ������IL-6��
������4���̼���ĸϸ����Ѫ��ϸ�������������⣬�˺���IL-9������EPOЭͬ��֧���������ϸ����ϸ��ϵ�ı����γɵ�λ��erythroid burst forming units,BFU-E���IJ�����IL-9������֧��BFU-E�Ķ��ڴ�
����IL-9������Hodgkin�ϲ��ķ����йء�ԭ����������Hodgkin������Reed-Sternbergϸ������IL-9��IL-9R��
������ʮ��IL-10
����1989������DNAX�о���Fiorentino�ȷ���С��Th2ϸ����D10.G4.1����һ���µ�ϸ�����ӣ�������Th1ϸ����ϸ������mRNA��ת¼����Ϊϸ�����Ӻϳ��������ӣ�cytokine synthesis inhibitory factor,CSIF��,ͬ������Ϊ��ϸ������10��interlenkin 10,IL-10����
����1.IL-10�IJ�������ԭ��˿��ԭ�̼�С��Th2ϸ����D10.G4.1�Լ�CDC25��CDC35��D9��MB2-1��ϸ���ɷ���IL-10����С�����ϸ��������ϸ��������ϸ����Ly1+��CD5+��������Bϸ��Ҳ�ɲ���IL-10�������࣬ijЩCD4+Tϸ����¡������AIDS����Bϸ��ϵ��EBV��Ⱦ���ܰ�ĸϸ����Burkitt���ܰ����������ϸ��������ѪTϸ��������CD8+ϸ����CD4+CD45RA+naive Tϸ����CD4+CD45RO+����Tϸ�������ɲ���IL-10��
����2.IL-10�ķ��ӽṹ�ͻ���С�����IL-10����λ�ڵ�1��Ⱦɫ�壬����������5�������Ӻ�4���ں��ӣ���������NF-��B��AP-1�Ľ��λ�á��˺�С��IL-10��DNA�Ͱ�����ˮƽ�Ϸֱ���81%��73%��ͬԴ�ԡ�DNA���з���������IL-10��EB�����������п��Ŷ�����I��BCRF-I����70%���ҵ�ͬԴ�ԡ����˰�BCRF-I�Ļ�������Ϊ����IL-10��vIL-10������ʾEBV������ȡ�˲��鶯��IL-10�Ļ����������������档��EBV��Ⱦ������ͨ������vIL-10��������ϸ��IFN-�ò���������EBV������ϸ��������ͷ�ֳ��Moore���ѿ�¡�ɹ�IL-10cDNA������COS7ϸ���еõ�����˺�С��IL-10����178��������л�������18�������ź���ϵ�У����ķ�����18.7kDa,PI8.1������IL-10����Ϊ160������л���С�����IL-10�����зֱ�5����4�����װ���л������ڲ�ͬ�ǻ�����ʹ���������������35��40kDa֮�䣬���������²��ȶ�������Һ�гʷǹ������ӵ�ͬԴ˫�塣��IL-10��������С��Դ��ϸ������С��IL-10���˵�ϸ���������á�
����3.IL-10������ѧ����
����(1)����С��Th1ϸ������ֳ�Լ�IL-2��IL-3��IFN-�á�TNF�Լ�GM-CSF��ϸ�����Ӻϳɡ������û���������IL-10������APCϸ����������MHC���ԭ�ı�����յ�APCϸ��������һ��ϸ�����ӣ��ı�ϸ�����źŵĴ���;�����Ӷ�ѡ��������ijЩϸ������mRNAת¼��IL-10��������TH0��TH-1��TH-2��Tϸ����¡����ֳ��
������2���ٽ��ʴ�ϸ��������ϸ����ֳ��IL-10Ҳ���ܰͽᡢƢ��ϸ�������ĸ������ӣ�cofac-tor)��
������3��ЭͬIL-2�յ�ConA�Ƣϸ����CTLǰ��ϸ����CTLp)�ֻ�Ϊ�����CTL��
������4�����Bϸ���Ĵ���ʣ��ٽ�Bϸ������ֳ��MHC���ԭ�����Լ�Ig�ķ��ڣ�����Th2��������IL-4��IL-5��Эͬ���á�
������5������NKϸ�����ӵIJ�����
����IL-10���������ܾ��п�EB���������ã���IL-10ͨ���ٽ�����ϸ������IL-1ra�����ܳ�Ϊ����֢�������ֶΣ�����ʵ�������IL-10����Ч�ر���LPS�յ�С���ݿ˶���ɵ�������
����(ʮһ��IL-11
����IL-11�����Paul�����鳤�ද��������ϸ����Pu-34�������巢�ֵġ������������ӿɴ̼�IL-6������С��ϸ����ϸ��ϵT1165.85.2.1����������ʹ�����кͻ��Կ�IL-6McAb�Ĵ��ڣ��������ִ̼����ã��Ժ�֤ʵ����������֬���γ��������ӣ�adipogenesis inhibitory factor,AGIF)��ͬһ���ʡ�1990������Ϊ��ϸ������11��interleukin 11,IL-11)��
����1.IL-11�IJ�������Ҫ�ɼ������Դ��ճ��ϸ����mesenchymal-derived adherent cell)��������������ϸ�������ʳ���άϸ�������߷γ���άϸ����������ϸ����IL-1�̼�Pu-34ϸ����IL-11�������ߡ�
����2.IL-11�ķ��ӽṹ�ͻ��������е����ȷ��ֲ���¡��IL-11��������С��IL-11������δ��¡�ɹ�����IL-11����λ��19��Ⱦɫ�壬����5�������Ӻ�4���ں��ӡ���5��UTR������RNA���ø���ϵ�TATA�У�3��UTR���ж��ATTTA�ظ����У������뽵��mRNA���ȶ����йأ����⣬����һЩ���п�����������յ�Ԫ���������ӣ���interferon-inducible elements��interferon-inducible factors)��SP-1��specificity protein 1 ��promotor-specific factor)��AP-1��activator protein 1)���ϡ��������IL-11���Ӻ�178��������л���������23kDa���������װ���л�������DZ�ڵ��ǻ���λ�㡣
����3.IL-11���塡����Ϊ�������Խ��IL-11����IL-11���������δ�����¡�ɹ���PU34ϸ��ÿ��ϸ����138��IL-11���λ�㣬����KdΪ1.2��10-10M��Ŀǰ�Ѿ�֤ʵ��gp130��IL-11������ź�ת���ǵ�λ��gp130��IL-11R�Լ�IL-6R��LIFR��OSMR��CNTFR�����еġ�IL-11���յ���ϸ���Ŀᰱ�����ữ��gp130�кͻ��Կ��������IL-11�յ��Ŀᰱ�����ữ���й�gp130�Ļ���ķ��ӵĽṹ�μ�����IL-6����ͱ��µ����ڡ�1994��Hilton�ȴӳ���С���cDNA�Ŀ��п�¡�ɹ�С��IL-11R����cDNA����IL-6R������24%ͬԴ��������Ѫ����������塣
����4.IL-11������ѧ���ԡ�IL-11�Ĺ�����IL-1��IL-6��G-CSF��SCF�Ĺ��������
������1���ٽ�Bϸ����������ɣ���һ����������CD4Tϸ���Ĵ��ڡ�����ʵ��֤����IL-11��ʹС��Ƣ�ԭ������PFCˮƽ��Ѫ���������Կ������ߡ�
������2���ٽ�ijЩIL-6����ϸ������TF-1��������IL-11��IL-6���յ���ϸ���������Ƶĵ����Ұ������ữ�Լ�ԭ������jun-B�ı��
������3����IL-3��IL-4Эͬ�����ڹ�����Ѫ��ϸ�������̸�ϸ��Go�ڡ�
������4����IL-3��Эͬ�ٽ������ϸ�����⼯���γɡ������ͳ��죬������ϸ���������������ѪѪС���������
������5��IL-11��С������̥��������Դ�IJ�ͬ�ֻ��κ�ϵ��ϸ�����д̼����ã������ڽ���Ҫ��IL-3��SCFЭͬ�����ڷֻ����ڣ�IL-11�����ɴٽ�CFU-E�ij��졣
������6���յ���ϸ�������ڵ��ϳɡ�
������7������֬����֬ø��lipoprotein lipase,LPL)���Ժ�֬��ϸ���ķֻ�����ˣ��ֳ�Ϊ֬���γ��������ӣ�AGIF����
����С������Ӧ��IL-11���յ��������ϸ���IJ�����ѪС�����ߣ����ӹ�����ԴCFU-GM��BFU-E��CFU-GEMM��ϸ��ϸ�����ڵ����ʡ��ڹ������Ƶ�С���У�IL-11�ɴٽ�����Ѫ��ѪС�塢��ϸ������ϸ�����������ڹ�����ֲС���У�IL-11�����Դٽ�����Ѫ������ϸ����ѪС��ˮƽ�Ļָ���
������ʮ����IL-12
����1982��Wagner�ȷ�����˿��ԭ�̼�С���ܰ�ϸ������������Һ�д���һ�ֲ�ͬ��IL-2��ϸ�����ӣ�����ϸ����������������IL-2Эͬ�ٽ���CTLӦ��1986�����˻���ܰ�ϸ��������MLC����PHA���PBMC����������Ҳ������������Ƶ����ӣ���ΪCTL�������ӣ�cytotoxic lymphocyte maturation factor,CLMF;��Tc maturation factor TcMF)��1991��Gubler�Ƚ�CLMf cDNA��¡������ɹ���������һ���µ�ϸ�����ӣ��콫CLMF����Ϊ��ϸ������12��interleukin 12,IL-12)��
����1.IL-12�IJ�������Ҫ��B�ܰ�ϸ��������
������1��MLC��˿��ԭ���PBMC�������塣
������2��PMA�����������A23187���ϴ̼�EBVת����B�ܰ���ĸϸ��RPMI8866�ɲ����ϸ�ˮƽ��IL-12��
����2.IL-12�ķ��ӽṹ�ͻ���IL-12���ɶ�������ӵ���Դ˫�壬�����ǵ�λ�ķ������ֱ�Ϊ35kDa��40kDa���ȵ����pH4.5��5.5���Ӹ߲�IL-12����B�ܰ���ĸϸ���ǿ�¡NC37.98�����Ŀ��п�¡��IL-12cDNA��תȾCOSϸ����ø߱����IL-12P35�ǵ�λ��197��������л�����7�����װ����3��N�ǻ���λ�㣬P40�ǵ�λ306��������л�����10�����װ��ᣬ4��N-�ǻ���λ�㡣С��IL-12P35��193��������л�������IL-12P35��66%ͬԴ�ԣ�P40��313��������л�����P40��70%ͬԴ�ԡ�����������£��ǵ�λ�еİ��װ���л�֮������γɸ��ӵķ��Ӽ������ṹ�������ǵ�λ���ɲ�ͬ�Ļ��������룬����תȾ������������ֻ�н����������ǵ�λcDNAͬʱתȾ���ܻ��������ѧ���Ե�IL-12��P35��IL-6��G-CSF��ͬԴ�ԣ�P40��IL-6���塢��״��Ӫ�����ӣ�CNTF�����塢G-CSF������ͬԴ�ԣ�����ϸ����������������������ʾIL-12���ӿ�����һ��ϸ������/������ϸ���������帴�����RPMI8866ϸ��ϵ�д���������Ȼɱ��ϸ���̼����ӣ�natural killer cell stimulating factor,NKSF)�Ľṹ������IL-12��ͬ��
���� 3.IL-12���塡IL-12R�ǵ�λ�Ļ��������¡�ɹ���������662�������ᣬ���ķ�����70kDa���ǻ�����ԼΪ100kDa��IL-12R�ǵ�λΪ���ʹ�Ĥ���ף���Ĥ������516��������л�����gp130��G-CSFR��LIFR�߶�ͬԴ������IL-12R�����IL-12��˫���Ѿ���IL-12R�ǵ�λ��IL-12��p40�ǵ�λΪ�������ϲ�λ�����Ϊ��������KdΪ2��6nM��PBMC������IL-12���������壬KdԼ100pM��Ŀǰ��IL-12R�ǵ�λ��ɸ���������һ���ǵ�λ��δȷ����IL-12P35�ǵ�λ��P40�ǵ�λ��IL-12R�ǵ�λ�ֱ���IL-6��IL-6R��gp130�кܸߵ�ͬԴ�ԣ��������ϵ�ģʽҲʮ�����ƣ���IL-12P35ͬP40�γ���Դ˫����ͬ˫���Ѿ����IL-12R�ǵ�λ��ϣ���IL-6ͬIL-6R�����Ϻ��ͬgp130�������ϡ�IL-12����ֲ���PHA���CD4+��CD8+Tϸ����Ⱥ�Լ�IL-2���CD56+NKϸ����1000��9000IL-12��ϵ�/ϸ����
����4.IL-12������ѧ���ܡ���IL-12���������������ԣ���IL-12��С��ϸ����������
������1����IL-2Эͬ�յ�CTL�ķֻ����ٽ�ͬ������CTL��Ӧ��
������2���̼�PHA�CD3+Tϸ��������CD4+��CD8+����ֳ��IL-12�յ�Tϸ�������ֳˮƽ����IL-2����ֳ�̼����ã����̼�50%�����ֳ����ϸ������Ũ��Զ����IL-2��IL-12������ֳ��������;����IL-2��IL-4��IL-7�Ĵ̼����ò�ͬ����Ϊ���IL-2��IL-2R��IL-4��IL-7�ĵ���¡���岻�����IL-12����ֳ���ã�IL-12����¡�����IL-2��IL-4��IL-7�յ���ֳ����������á�IL-12�Ծ�ֹPBMC��������ֳ���ã���һ������IL-4���ơ���IL-12�����ʼ���IL-2��Эͬ�յ���ֹPBMC��ֳ������������ǣ���IL-12�ٽ�IL-2�յ�Tϸ��IL-2r P55�ı�����PBMC��IL-2�ķ�Ӧ�ԣ���IL-2�ٽ�PBMCijЩ��ȺIL-12R�������IL-2���ڵ������£�IL-12�����IFN-��mRNAת¼ˮƽ���������ȶ��ԣ��ٽ�����IFN-�ã���С��IL-12���յ�Th1ϸ������IFN-�á����⣬IL-12ͨ���յ�������IFN-�ü������ϸ�����յ���TNF-���ķ��ڡ�IL-12���յ�Th1��Ⱥ���γɣ������ÿ���ͨ������;������IL-12ֱ��������Th1��Ⱥ����IL-12�̼�Tϸ����NKϸ������IFN-���յ�Th1��Ⱥ�γɡ�
������3��ЭͬIL-2�յ�CD56+NKϸ����ֳ�Լ�LAKϸ���������ٽ�ADCC���ܣ�NKSF�ͷţ��յ�CD56��CD2��CD11a��IL-2R��TNFR��CD120b����LFA-1��ICAM-1�ȷ��ӵı��
������4���ٽ�Bϸ��Ig������Ig����ת������IgMתΪIgG������IL-4�յ�Bϸ��IgE�ϳɡ���������������Tϸ�������ģ������û�����IFN-�á�TGF-�º�IL-8����IgE�����Ļ�������������ͬ��
����IL-12��������ѧЧӦ�����ϸ������Ũ�Ⱥܵͣ���pM���������ʼ���IL-2����Ӧ�ÿɽ���IL-2������ͬʱ���CTL��NK��LAK��ɱ�˻��ԣ����IL-12���ܳ�Ϊһ���µĿ����������Ƽ���
������ʮ����IL-13
����1993��Minty�ȱ����˰�ϸ������12��interleukin 13��IL-13��cDNA��¡��óɹ���IL-13��Ҫ�ɻTϸ����������CD28������յ�IL-13mRNA�����С��IL-13��Th2��Ⱥ������
����1.IL-13�ķ��ӽṹ�ͻ�����IL-13����λ��5��Ⱦɫ�壬��4�������Ӻ�3���ں�����ɡ��˺�С��IL-13�ڻ���ˮƽ����66%ͬԴ�ԡ���IL-13������132�������ᣬ�г�18����������ź��ĺ���IL-13����Ϊ114�������ᣬ���ǻ���IL-13������Ϊ12.4kDa���ǻ�����Ϊ17kDa����С��IL-13��58%ͬԴ�ԡ�IL-13������IL-4�����������ڰ�����ˮƽ����20%��25%ͬԴ�ԡ�
����2.IL-13������ѧ����
������1����������ϸ�����ӳ�����ϸ����������ʱ�䣬����LPS�յ�����ϸ��������ϸ��IL-1��IL-6��IL-8��TNF-������֢���Ӳ�����
������2��Эͬ��IgM�Bϸ������ֳ���յ����ϵ�Bϸ��MHC���ԭ��CD23��CD72�ı���յ�Bϸ������IgM��IgG��IgE��
������3���յ�������ܰ�ϸ����LGL������IFN-�ã�������IL-2Эͬ�̼�LGL����IFN-�ã�������յ�LAK�����Լ�Th1��ϸ�������п�������Ҫ���á�
������ʮ�ģ�IL-14
������ϸ������14��interleukin 14��IL-14���ֳƸ߷�����Bϸ���������ӣ�high molecular-weight B cell growth factor,HMW-BCGF)������IL-14��468�������ᣬ������ʽ����Ҫ��Tϸ�����������յ��Bϸ������ֳ�����Ծ�ֹBϸ���̼����ã�������˿����ԭ�̼���Bϸ��Ig�ķ��ڡ�
������ʮ�壩IL-15
����Grabstein�������ں�����Ƥϸ��ϵCV-1/EBVA�����з���һ����ά��CTLL��ֳ�����ӣ��������¡��óɹ�������Ϊ��ϸ������15��interleukin 15,IL-15)���Ժ�IL-15 cDNAΪ̽���¡��IL-15 cDNA��óɹ����˺ͺ�IL-15���������������97%ͬԴ�����������֯��ϸ������IL-15mRNA�����ġ��Ρ��Ρ���������̥�̺������Լ�PBMC��Ϊ�ḻ�����������ѪTϸ��ȴ��ⲻ��IL-15mRNA��IL-15ǰ�������162��������ɣ��ȵ����нϳ�Ϊ48��������л����������114��������л���������14��15kDa��IL-15��������IL-2�Ľṹ���ܣ��̼�CTLLϸ����PHA�Tϸ����CD4+CD8+��Ⱥ������ֳ���յ�CTL��LAKϸ���IJ�����IL-15�Ĵ̼�������Ҫ�а�ϸ����IL-2����ºͦ����IJ��룬������ҪIL-2����Ħ�����
��4-3����ϸ�����أ�IL���������Ҫ����ѧ����
| IL | ��Ҫ����ϸ�� | ��������Ŀ | ��������kDa) | ��Ҫ����ѧ���� |
| IL-1 | ����ϸ�� | IL-1��159 | 17.5 | ��1���ٽ�����ϸ����Tϸ�������ֳ�ͷֻ� |
| �� | ����ϸ�� | IL-1��153 | 17.5 | ��2����ǿCTL��NKɱ�˻��� |
| �� | �� | �� | �� | ��3��ЭͬIL-4�̼�Bϸ����ֳ���ֻ���Ig���� |
| �� | �� | �� | �� | ��4��ЭͬCSF�ٽ���Ѫ���� |
| �� | �� | �� | �� | ��5���̼���ϸ������SCF |
| IL-2 | �Tϸ�� | 133 | 15.5 | ��1���ٽ�� Tϸ����ֳ���ֻ���ϸ�����Ӳ��� |
| �� | �� | �� | �� | ��2����ǿCTL��NK��LAKɱ�˻��� |
| �� | �� | �� | �� | ��3���ٽ�Bϸ����ֳ���ֻ��Ϳ������ |
| �� | �� | �� | �� | ��4�������ϸ�� |
| IL-3 | �Tϸ�� | 133 | 15 | ��1���ٽ����ܸ�ϸ����������ϸ�����������������ˡ�ǰ�������ˡ����ԡ����ᡢ�ʴ�ϸ����ֳ�ͷֻ� |
| �� | �� | �� | �� | ��2���ٽ�Tϸ����ֳ |
| �� | �� | �� | �� | ��3����ǿ����ѪPMN��Mo��Eo���������ٽ�Eo��ADCC |
| IL-4 | �Tϸ����Th2) | 129 | 20 | ��1���ٽ��Bϸ����ֳ��Ig���������ת��ΪIgE��IgG1 |
| �� | �� | �� | �� | ��2��Tϸ���������� |
| �� | �� | �� | �� | ��3���ٽ��ʴ�ϸ����ֳ |
| �� | �� | �� | �� | ��4����ǿ����ϸ������ |
| �� | �� | �� | �� | ��5��ЭͬCSF�̼���Ѫϸ�� |
| IL-5 | �Tϸ�� | 115��2 | 45 | ��1���յ�Bϸ���ֻ�Ϊ�������ϸ����IgA�ϳ� |
| �� | ��Th2) | �� | �� | ��2��ЭͬIL-2�ٽ�CTL�ֻ� |
| �� | �� | �� | �� | ��3���ٽ���Eo�����ֳ�ͷֻ� |
| IL-6 | ����ϸ�� ����ϸ�� |
184 | 26 | ��1���̼�Tϸ����Bϸ�����ӽ���ϸ����ϸ����ֳ |
| �� | ����άϸ�� | �� | �� | ��2���ٽ�Bϸ��Ig���� |
| �� | �� | �� | �� | ��3���ٽ�CTL��NK��ϸ���;�ϸ���ֻ� |
| IL-7 | ��������� | 152 | 25 | ��1���̼�Bϸ��ǰ�塢ǰBϸ������ֳ |
| �� | �л���ϸ�� | �� | �� | ��2���ٽ�ǰTϸ��������ϸ����Tϸ����ֳ |
| �� | �� | �� | �� | ��3���յ�LAK���� |
| IL-8 | ����ϸ�� | 69,72,77, | 8-10 | �������ԡ��ȼ�����ϸ����Tϸ�� |
| �� | ��Ƥϸ�� | 79 | �� | |
| IL-9 | �Tϸ�� | �� | 14.2 | ��1��Tϸ���������� |
| �� | 126 | �� | ��2��ЭͬIL-3�̼��ʴ�ϸ�� | |
| IL-10 | �Tϸ�� | 2��160 | 35��40 | ��1������Th1ϸ������ϸ������ |
| �� | ��Th2) | �� | �� | ��2���ٽ�����ϸ���ͷʴ�ϸ����ֳ |
| �� | �� | �� | �� | ��3��ЭͬIL-2�ٽ�CTL�ֻ� |
| IL-11 | ������ά�� | 178 | 23 | ��1���ٽ�Bϸ��������� |
| �� | ϸ�� | �� | �� | ��2���ٽ���ϸ�������� |
| �� | �� | �� | �� | ��3��ЭͬIL-3�ȴ̼������ϸ���;�ϸ������ |
| IL-12 | Bϸ�� | 197/306 | 35/40 | ��1��ЭͬIL-2�ٽ�CTL��NK��LAK�ֻ� |
| �� | �� | �� | �� | ��2���ٽ�PHA�Tϸ����ֳ |
| �� | �� | �� | �� | ��3���ٽ�Bϸ��Ig����������ת�� |
| IL-13 | �Tϸ�� | 132 | 17 | ��1���̼�Bϸ����ֳ��CD23���� |
| �� | �� | �� | �� | ��2�����Ƶ���-����ϸ����֢��ϸ�����Ӳ��� |
| IL-14 | �Tϸ�� | 468 | - | ��1���̼��Bϸ����ֳ |
| �� | �� | �� | �� | ��2������˿��ԭ�յ���Bϸ��Ig�ķ��� |