四、免疫球蛋白的多样性
机体对外界环境中众多抗原刺激可产生相应的特异性抗体,有人推算抗体的多样性在107以上,这种抗体多样性主要是由遗传控制的。引起免疫球蛋白多样性的原因主要有以下几个方面。
1.胚系中众多的V、D、J基因片段 胚系(germ line)中未重排的(unrearranged) DNA有众多的V基因片段以及一定数量的D、J基因片段。以小鼠为例。VH、DH和JH基因片段分别为1000、12和4,单独重链重组的多样性可达4.8×104左右;Vκ和Jκ分别约为250和4,κ轻链V-J重排的多样性为1.0×103。经重链与κ轻链随机配对后推算的多样性为(4.8×104)×(1.0×103)=4.8×107。
表3-5 小鼠Ig胚系基因片段和重链、κ轻链配对的多样性
| 多肽链 | 基因片段数 | 可变区基因 | 经重排和随机配对后 | |
| V J | 重组方式 | 推算的多样性数目 | ||
| 重 链 | 1000 12 4 | V-D-J | 4.8×104 | |
| 4.8×107 | ||||
| κ轻链 | 250 | V-J | 1.0×103 |
注:多样性数目不包括VDJ连接多样性、N区插入和体细胞突变所增加的多样性数目。
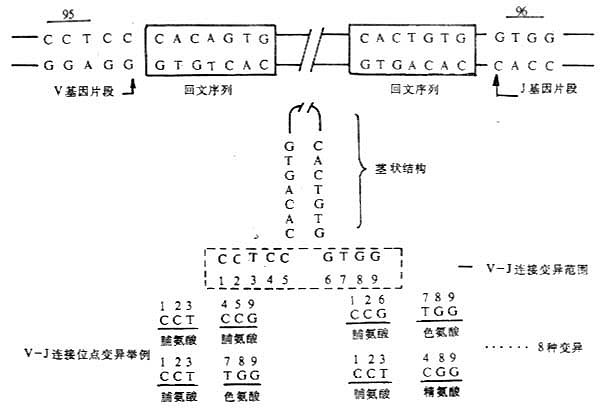
2.VDJ连接的多样性 轻链基因重排过程中,VJ连接点以及重链基因重排过程中D-J以及V-D-J连接点有一定的变异范围。例如轻链VL因片段3′端5个核苷酸CCTCC和JL基因片段5′端4个核苷酸GTGG连接时,9个核苷酸中只有6个核苷酸编码轻链第95、96位氨基酸,可产生8种不同的连接方式(图3-13)。
3.体细胞突变(somatic mutation) 体细胞在发育过程中可发生基因的突变。以长期体外培养的B细胞前体为例,每个细胞每个碱基对的突变率为1~3×10-5,这种类型突变主要发生在V基因。体细胞突变扩展了原有胚系众多基因片段的多样性。
4.N区的插入(insert of N region) 在Ig重链基因片段重排过程中,有时可通过无模板指导的机理(non-temple directed mechanism)在重组后D基因片段的两侧即VH-DH或DH-JH连接处插入称之为N区的几个核苷酸。N区不是由胚系基因所编码。在N区插入前,先通过外切酶切除VH-DH或DH-JH连接处的儿个碱基对,然后通过末端脱氧核苷酸转移酶(terminal deoxynucleotidyl transferase,TdT)连接上N区。由于额外插入了N区,可发生移码突变(frameshift mutation),使插入部位以及下游密码子发生改变,从而编码不同的氨基酸,增加了抗体的多样性。
图3-13 轻链基因V-J连接多样性举例
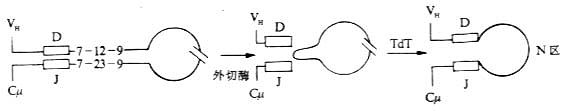
图3-14 N区插入D-J连的接处(示意图)
5.轻链重链相互随机配对 如表3-5所示,小鼠重链与κ轻链随机配对后推算的多样性可达4.8×107,如果再加上重链与λ轻链的随机配对其多样性就更多了。