三、免疫球蛋白基因表达的调节
(一)核因子对Ig基因转录的调节作用
Ig基因转录活性是由两个顺式作用元件(cis-acting elements)即启动子(promoter,P)和增强子(enhancer,E)来调节。而启动子和增强子的功能又由反式作用的核因子(trans-acting nuclear factor)所控制。这些核因子也称为DNA结合蛋白(DNA-binding proteins,DBP),结合到启动子或增强子内特异的核苷酸序列,从而抑制或刺激启动子和增强子的活性。许多种类细胞受到某些刺激后可诱导或活化特定的DBP。
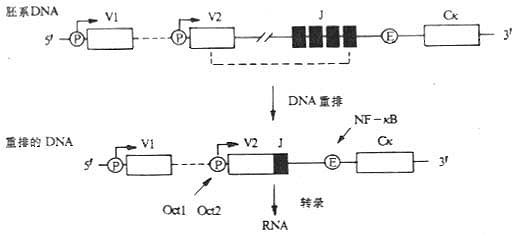
1.ATGCAAAT序列及其核因子 Ig重链启动子含有8个保守的核苷酸序列ATGCAAAT,这个序列也存在于κ轻链的启动子、重链增强子以及许多非Ig的启动子中。B细胞Ig基因的转录依赖于启动子中这一完整的ATGCAAAT序列的存在。哺乳动物细胞中至少存在三种特异性结合ATGCAAAT的蛋白质,即Oct1、Oct2(OTF2A)和OTF2B。Oct1又称OTF1(octamer transcription factor 1),广泛存在于哺乳动物的细胞中;OTF2A和OTF2B为淋巴细胞特异性,活化Ig基因的转录。
2.GGGACTTTCC序列和NF-κB 小鼠κ链增强子含有GGGACTTTCC10bp 的序列,是一种称之为NF-κB(nuclear factor of kappa B)DNA结合蛋白结合的靶序列。
(1)NF-κB结合靶序列的分布:NF-κB结合的DNA序列也存在于许多其它淋巴细胞、非淋巴细胞、病毒基因增强子和启动子以及HIV-1的长未端重复序列中,如某些MHCⅠ类基因,IL-2Rα链,TCRβ链,IL-2、IL-3、IL-6、IFN-α、IFN-β、IFN调节因子、GM-CSF、SV40基因等。在不转录κ基因的前B细胞(pre-b cell)系中无NF-κB活性,这些细胞用LPS处理后可诱导出有功能的NF-κB,随之也刺激κ链基因的转录。
(2)NF-κB结构及作用特点:NF-κB是一个异源四聚体复合物,包含两个与DNA结合的50kDa多肽和两个不与DNA直接结合的65kDa多肽链NF-κB解离常数在10-12~10-13M之间,这种高亲和力是它发挥功能所必需的。在大多数细胞系中由于抑制因子IκB与p65的偶联作用,使整个NF-κB分子以无活性的形式存在于细胞质内。IκB由于磷酸化(如PKC作用)或其它的修饰作用丧失功能,从NF-κB/IκB复合物上解离下来,洲离的NF-κB随即进入细胞核,结合靶基因的特定序列,从而激活这一基因的转录反应。如抗原结合到B细胞mIg后刺激B细胞内PKC活化。LPS也可活化PKC。NF-κB在有效合成κ链的骨髓瘤细胞中是无活性或是缺乏的,提示NF-κB和κ链基因的增强子在B细胞分化早期对于起始κ基因转录是需要的,但对于分化后期维持转录过程中并不是必需的。
图3-12 免疫球蛋白基因转录的调节
注:VDJ重排使启动子(P)靠近位于Jκ和Cκ之间的增强子,增加了已重排V2基因的转录水平,DNA结合蛋白NF-κB结合到增强子内靶序列,Oct1、Oct2可结合到启动子上。
(二)等位基因排斥现象
免疫球蛋白基因表达中一个重要的发现是等位基因排斥现象(allelic exclusion)。编码人Ig重链基因位于第14对染色体上,在B细胞发育过程中,这一对同源染色体中仅有一条染色体Ig基因得到表达。例如一个人B细胞内来自母本的染色体上的Ig重链基因得到表达,则来自父本的第14号染色体的Ig重链基因受到抑制而不表达。这可能是通过已发生有效重排的染色体产生反馈抑制(feedback inhibition)信号抑制另一条同源染色体上的Ig重链基因发生重排。一条染色体上Ig重链基因的有效重排,一方面抑制了另一条同源染色体重链基因的重排,另一方面可发出信号使第2号染色体上Igκ轻链基因发生重排,如果一对同源染色体上的κ轻链基因重排均无效,才发生22号染色体λ轻链基因重排,这种现象称为轻链同种型排斥现象(light chain isotype exclusion)。