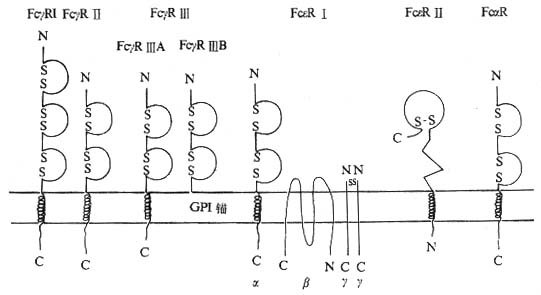
��������������Fc������
����Ig������������ԭ�ԵIJ����ΪIgM��IgG��IgA��IgD��IgE���࣬����Ig�IJ�ͬ������Ҫ����ṹ�йء�����������ϸ��������в�ͬ��Ig Fc�����壬ͨ��Fc������Ig Fc�Ľ�ϣ�����Ig�鵼���������ܻ������˹��̡�Ŀǰ�Ѽ�����ȷ����CD��ԭ��Fc������Fc��R��Fc��R��Fc��R��
������һ��Fc��R��CD64��CD32��CD16��
����1.Fc��R�Ľṹ�ͷֲ���Fc��R�ɷ�ΪFc��R��Fc��R���Fc��R�����࣬���ǵĽṹ�ͷֲ�������ͬ��
������1��Fc��R��CD64����70kDa��Ĥ�ǵ��ף���Ig�������Ա����Ĥ������3��C2�ṹ������Ⱦɫ�嶨λ��1q23��24��ʶ��CD64�Ĵ�����McAb��McAb22��McAb32.2��197��10.1��Fc��R���Ǹ��������壬KdֵΪ10-8��10-9M����Ҫ���˵ĵ���IgG1��IgG3�Լ�С��IgG2a��IgG3��ϡ�����IgG4��ϵ�������ȷ���ͣ���IgG2�����������Fc��R����Ҫ�ֲ��ڵ���ϸ��������ϸ����������ϸ���ȣ�������ˮƽ������ͬ��Fc��R��λ������15000��40000/ÿ������ϸ����>50000/����ϸ����<1000/����������ϸ����IFN-�ÿɴ̼�����ϸ��������ϸ����������ϸ������Fc��R��ˮƽ����5��10����G-CSFҲ�����ִٽ����á�
������2��Fc��R��CD32����40kDa��Ĥ�ǵ��ף�����Ig�������Ա����Ĥ������2��C2�ṹ������Ⱦɫ�嶨λ��1q23��24��ʶ��CD32�Ĵ�����McAb��CIkM5��IV��3��KuFc79��41H16�ȡ�Fc��R���뵥����IgG1��IgG3��IgG4���Ϊ��������Kd5��10-7M��Fc��R������ڳ���ϸ���������Ѫϸ����������Ŀ��20000��40000/ÿ��ϸ��������DNA���к��ܲ�ͬ��Fc��R��ɷ�Ϊ������ʽ����ͬ��ʽFc��R��IJ����Ҫ���ڰ������Ľṹ��ͬ��
������3��Fc��R��CD16����50��70kDa�ǵ��ף���Ig�������Ա����2��C2�ṹ������Ⱦɫ��λ��1q23��24��ʶ��CD16�����Ե�McAb��HUNK2��Leu11��3G8��Gran1��B73.1�ȡ�Fc��R������IgG��IgG3��Ϊ���������塣Fc��R����Fc��R��A��Fc��R��B�������ͣ���Fc��R��A����Ĥ�ṹ����Ҫ�ֲ��ھ���ϸ����NKϸ������������ϸ��������ϸ�������ˮƽFc��R��A��������ϸ������ˮƽ�ϵ͡�Fc��R��A���������ӵ�CD3�ƻ�Fc��R�����˫����أ�����ϸ����Fc��R��A��CD3�����������أ�NK/LGL��Fc��R��A���������ء�TGF-�´ٽ������ĵ���ϸ������Fc��R��A����Fc��R��B��ͨ��GPI��ê����������ϸ�����棬ÿ����������ϸ������10��20��ѪҺ�п����Ե�Fc��R����Ҫ����������ʽ��������ϸ���������ʱ�䴦��������Խ���Fc��R��B�ı���ˮƽ��������ͨ��������Դ�Ե���ø�г�GPI���ӷ����йء�
ͼ1-8 Fc��R��Fc��R��Fc��R�ṹʾ��ͼ
����2.Fc��R�Ĺ��ܡ�Fc��R�Ĺ�����Ҫ��ͨ������ϸ����NKϸ�������ӵġ�
������1������-����ϸ����Fc��R��͢���ɽ鵼�˵���ϸ��ADCC��ɱ�������Ȱ�ϸ��������ADCCЧӦΪMg2+����������LFA-1��ճ�����Ӳ��롣IFN-�ÿɴٽ�����ϸ��Fc��R��鵼��ɱ�����á�����-����ϸ����ͨ��Fc��R�ӵ������ɺ�������߸���������á�
������2��������ϸ�������ʷ����������ϸ������ͨ��Fc��R�ܽ��ϸ��������IFN-�ô̼��¿�ͨ��Fc��R���Fc��R��鵼ɱ�����ã�����Fc��R��IFN-����Ҫ���յ������ˮƽ���ߣ�����Fc��R�����ˮƽ��δ���ı䣬������ͨ����ɱ�˻����ĵ��ڡ�GM-CSFҲ��ͨ��Fc��R����ȷ����������ϸ����ɱ��ˮƽ��GPI���ӵ�Fc��R��B�����ܽ鵼������ϸ��ɱ�����������á��������ϸ��ͨ��Fc��R�ӵ������ɺ�������߸���������á�
������3����������ϸ����δ�̼�����������ϸ��û��ɱ�����ã�GM-CSF��TNF��IL-5������������ϸ������ADCCЧӦ����Ч���������ɱ�˼�����Ϳ�����������Ҫ���á�GM-CSF����������ϸ���ļ���������Ҫ��ͨ��Fc��R��鵼�ġ�
������4��NKϸ����ͨ��Fc��R��A�鵼ADCCɱ������ϸ���Ȱ�ϸ����IL-2��IFN-�ÿ��������NKϸ����ɱ�˻��ԣ����������Ըı�Fc��R��A�ı���ˮƽ��
����������Fc��R��CD89��
����Fc��R��CD89��Ϊ������60kDa��Ĥ�ǵ��ף���Ĥ����206�����ᣬ��Ĥ��19�����ᣬ������Ϊ41�����ᣬ��Ig�������Ա����Ĥ����2��C2�ṹ��Ϊ���������壬��Ҫ�����ڵ���ϸ��������ϸ����������ϸ���ȣ��鵼���ɡ�ADCC�Լ���֢���ʵ��ͷš�������ϸ������Fc��R�ɽ��Ѫ���ͺͷ�����IgA1��IgA2������ԼΪKd5��10-7M���Ȼ�ѧ����������IgA�ɴ̼�������ϸ���ѿ�����
����������Fc��R��Fc��R��Fc��R��
����1.Fc��R�Ľṹ�ͷֲ� Fc��R�ɷ�ΪFc��R���Fc��R�����࣬��ṹ���ֲ����鵼������ѧ����������ͬ��
������1��Fc��R��Ϊ���������壬Kd10-9��10-10M���ɦ����¡���-����������ɡ����Ц�����222��������л���������Ϊ25kDa����Ĥ������Ig������ṹ��2��C2������Fc��R���Fc��R��߶�ͬԴ����Ĥ������IgEC��2/C��3��ϣ���Ĥ��20���Ұ������к�����Fc��R��A��ͬ��8��������л�����������31��������ṹ��Ϊ���ء�������243��������л���������Ϊ33kDa�����ĸ���Ĥ���֣�N����C�˶�λ�ڰ����ڣ��������ܰѦ����ͦ�-��������һ�����������ɶ�����������ͬԴ�����壬ÿ����62�������ᣬ������Ϊ8kDa����Ĥ����ֻ��5��������л���������CD35�߶�ͬԴ��������Fc��R�������ȶ��Ժ��źŵ�ת���йء�NKϸ������Fc��R��A��CD16��������CD3�ƻ�Fc��R�������������ʾFc��R�������CD3�������ЦƵĽṹ���ܵ������ԡ� Fc��R����Ҫ�ֲ����ȼ�����ϸ���ͷʴ�ϸ����
������2��Fc��R��CD23�������������壬������45kDa��������Ĥ�ǵ��ף����Ϳ�Ĥ���ף���C��ֲ��Ѫ���ؼ����Ա��CD23����321�������ᣬN���ڰ�Ĥ�ڣ�1��23λ��������ɰ���β��24��43λ������Ϊ��ˮ��Ĥ������C�˰�Ĥ������277����������ɣ���һ���ǻ����㣬82��102��125��150λ������л�Ϊ����ˮ��ø����λ�㣬������ͬԴ��λ��C��163Cys��282Cys֮�䣬��ͬԴ������6��Cys��88��116λ������֮����һ�������������ṹ������CD23����ͬԴ��������γɡ�CD23���ӿ���Ĥ��C���ѽ�IJ�ͬƬ��14kDa��25kDa��33��37kDaƬ�ξ���ΪIgE������ӣ�IgE-binding factor IgE-BF����Fc��R����ڵ���ˮ��ø�ѽ���γɿ�����CD23���ӣ�sCD23����IgE-BF��CD23mRNA��Fc��R��amRNA��Fc��R��bmRNA���֣������������CD23���ӽ��ڰ�������7��������л��IJ��ͼ1-9����Fc��R��a��Bϸ�����������ΪsCD23��Fc��R��b������Bϸ����Tϸ������������ϸ����ѪС�塢����ϸ��������ϸ������ͻ״ϸ�����ʺ���ϸ��������EBV������ı��ʰ�ϸ��������ϸ��ϵ��U937�ȣ���Ҫ��Ĥ������ʽ���ڣ�IgEC��3��Fc��R�����йء�IL-4���յ�����Bϸ��������ϸ������������ϸ��ת¼Fc��R��bmRNA���ٽ�CD23�ĺϳ�����EBV�˵���EBNA2��CD23���PsCD23���ͷ�Ҳ�дٽ����á���IFN-�á�TGF-�¡�PGE2����Ƥ�ʼ��صȶ�Bϸ������CD23���ͷ�sCD23�����������á�
����2.Fc��R����
������1��Fc��R���ȼ�����ϸ���ͷʴ�ϸ�����и�����Fc��R��ÿ��ϸ������Լ��10���������Ӧ��Ӧԭ�놨������ϸ�����ʴ�ϸ������IgE/Fc��R�����Ϻ�ͨ������ʹ���ἡ��ˮ�⣬����Ca2+Ũ�����ӣ�ʹϸ���ѿ������ϳɺ��ͷ���֯����LT��PAF�ȶ��ֽ��ʣ��鵼�����ٷ��ͳ�����Ӧ��
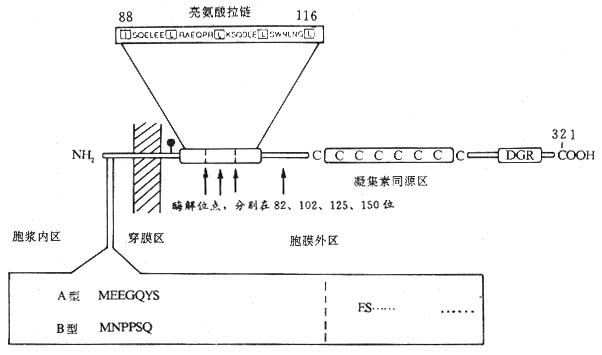
ͼ1-9 CD23���ӽṹģʽͼ
������2��Fc��R��Fc��R��ΪBϸ���������ԭ����̬��Ӧ�Լ�������PBMC��CD23�ܶ��������ӣ�Ѫ��IgE-BP(sCD23�����ߡ�sCD23����Bϸ���������ӣ�b cell growth. factor, BCGF)���ԣ����ֳ�Bϸ����Դ��Bϸ���������ӣ�B-BCGF����sCD23�����������������ÿ�������Ϊ����������CD21��CR2����Ϻ�鵼�ģ�CD23����ͨ���ɽ��̼ˮ�������������ͬԴ����CD21������ϡ����⣬sCD23ͨ�������������ṹ������Bϸ��ĤCD21���ӽ������ٽ�Bϸ��������sCD23��ĤCD23�����������ã��ٽ�Bϸ���ķֻ���IgE�IJ���������IL-4��Эͬ���ã����⣬Fc��R�ɽ鵼IgE������ADCC���������á�CD23��B�ܰ�ϸ����ת��������йأ�EBVת��Bϸ����ֻ��Bϸ������CD23���ܽ�����������ϸ��ϵ��������EBV�˵���EBNA2�йأ�EBNA2�����Fc��R��a������ʼ��λ-275��-89��һ��189bp DNAƬ�ν�ϣ���ʽ������Fc��R��a���������ӣ����յ�Bϸ���߱���CD23��sCD23����ĤCD21���ӽ������γ�һ���Է����������ơ�����B�ܰ�ϸ����Ѫ����B-CLL������Bϸ������CD23���ӣ�����Ѫ����sCD23ˮƽ�������ߡ�
��������IgM���塢IgD����Ҳ�б�����ǰ����Ҫ������Bϸ�������߱����ڳ���Bϸ����������Ig Fc������δ����CD�ı�ţ����ڱ��ڴ��ԡ���������IgA��IgM��Ĥת�����������Һ���йصĶ��Ig���壨Poly-IgR),ͨ������Ig�е�J����϶��鵼Igת�˹��ܣ�Poly IgR�������׳������Ա��
����Ȫ��
�����
����1.Arnaout MA.Structure and function of the leukocyte adhesion molecule CD11/CD18.Blood,1991;75(5):1037
����2.Autran B,et al. T cell receptor ��/��+lymphocyte subsets during HIV infection. Clin Exp Innunol,1989;75:206
����3.Barber EK,et al.Tthe CD4 and CD8 antigens are coupled to a protein tyrosine kinase(p56lck)that phosphorylates the CD3complex.PNAS,1989;86:3277
����4.Bevilacqua MP,et al.Endothelial leucocyte adhesion molceule 1,An inducible receptor for neutrophils related to complement regulatory proteins and lectins.Science,1989;243:1160
����5.Carel JM,et al.Dual antigen recognition by B cells,Immunol Today,1993;14(1):8
����6.Eiden LE,et al.HIV interactions with CD4:continum of conformations and consequences. Immunol Today,1992;13(6):291
����7.Figdor CG,et al.On the mode of action of LFA-1. Immunol Today,1990;11(8):277
����8.Hirsch R,et al.Effects of in vivo administration of anti-CD3 monoclonal antibody on T cell function in mice. j Immunol,1989;142(3):737
����9.Hogg N.The leukocyte integrins. Immunol Today,1989;10(4):111
����10.Holoshitz J,et al.Isolation of CD4 CD8mycobacteria-reactive T lymphocyte clones from rheumatoid arthritis synovial fluid. Nature,1989;339:226
����11.Kavanough AF,et al.Role of CD18 in adhesion and transendotlial migration of T cell. J Immunol,1991;146(12):4149
����12.Kevin H,et al.Murine hematopoietic cell surface antigen expression. Immunol Today,1988;9(11):344
����13.Knapp W,et al.Towards a better definition of human leucocyte surface molecules. Immunol Today,1989;10(8)253
����14.Littman DR,et al.The isolation and sequence of the gene encoding T8, A molecule defining functional classes of T lymphocytes. Cell,198540(1):237
����15.Marx J.Biologists turn on to ��off-enzymes��.Science,1991;251:744
����16.Modlin RL,et al.Lymphocytes bearing antigen-specific T-cell receptors accumulate in human infection disease lesions .Nature,1989;339:544
����17.O'Brien RL,et al.Stimulation of a major subset of lymphocytes expressing T cell receptor �æ�by an antigen derived from mycobacterium tuberculosis. Cell,1989;57:667
����18.Patel MD,et al.Multiple Kinases and signal transduction.JBC,1987;262(12):5831
����19.Pinto A,et al.New molecules burst at the leukocyte surface A comprehensive review based on the 5th international workshop on leukocyte differentiation antigens Boston, USA,3-7November,1993.Leukemia,1994;8(3):347
����20.Rivas A,et al.CD4 molecules are associated with the antigen receptor complex on activated but not resting T cells. j Immunol,1988;140(9):2912
����21.Rudd CE.CD4 CD8 and the TCR-CD3 complex:a novel class of protein tyrosine kinase receptor. Immunol Today,1990;11(11):400
����22.Sanders ME,et al.Human naive and memory t cells:reinterpretation of helper-inducer and suppressor-inducer subsets. Immunol Today,1988;9(7):195
����23.Schlossman SF,et al.Leucocyte typing V:white cell differentiation antigens. Oxford University Press, Oxford 1994 in press
����24.Sukhatme VP,et al.Gene for the human T cell differentiation antigen Leu-2/T8 is closely linked to the light chain locus on chronosome 2. J Exp. Med,1985;161(2):429
����25.Williames AF.A year in the life of the immunoglobulin superfamily. Immunol Today ,1987;8(10):298
����26.��谣���³ϣ����������������������ϸ���ֻ���ԭ������.����¡����ͨѶ��1994;10(3):70