
�ڶ��ڡ�ѪҺ����
����ѪҺ�Ŀ��������ʶԻ�������Ҫ�������á���Ѫ��ϵͳ����ʱ������Ѹ�ٿɿ��ط���������Ծ����ܼ��ٳ�Ѫ��ѪС�����(ճ�Ա�̬)���ڷ�����ã����ַ������Ҫ����ά�����������֧�֣������ߵ��γ��Ƕ�����Ѫ��������ã�����һϵ��ø�ٷ�Ӧ�Ľ����Ŀǰ�ѷ��ֵ���Ѫ������14��(��10-3)��
������Щ��Ѫ���ӳ�Ca2�����Ϊ�����ʣ�������ɸ���ϳɵ�Ѫ���ǵ��ף����Ǵ��������ˮ��ø�ࡣ��7��Ϊ����øԭ������Ѫ�����б�������Ե���Ѫ���������������ʾ����������Ը��ӵ��½���a������ʾ��
ͼ10-1��ѪҺ���̹���
����һ��ѪҺ���̵Ļ���
����ѪҺ���̵Ļ�ѧ�������ܽ�״̬����ά����ԭת��Ϊ����״̬����ά���ף����˷�Ӧ������Ҫ����Ѫø��������ѪҺ�������Ե���Ѫøԭ��ʽ���ڣ���һ�������±��������Ϊ��Ѫø����Ѫøԭ���������ɻ����Ѫ��������֬�������γɵĸ������ˣ���Ѫ���ӵĻ�ǵ���ѪҺ���̵Ĵ������ơ��ݴ�����Ѫ���̵ķ�ʽ��ͬ��������Դ��(intrinsic)����Դ��(extrinsic)��Ѫ֮�֡���Դ����Ѫָ����Ѫ����Ĥ�����ѪҺ�������Ӵ��������������ģ�����Ѫ������Ѫ���Ӳ������Ѫ���̣�����Դ����Ѫ��ָ��������֯�ͷŵ���֯��Ѫ�������������Ѫ���̡�ѪҺ���̹��̵Ĺ��ſ�ͼ�����£�(ͼ10��1)
����������Ѫøԭ�����������
������Ѫøԭ�������ɻ����Ѫ����Xa��Va��Ca��������֬�������ɵĸ����塣����X������ΪXa�Ǵ˹��̵Ĺؼ����衣����X�ļ���������;��������Դ�Ժ���Դ��;����
��10-3����Ѫ�����������䲿������
| ��Ѫ���� | ͬ���� | �ϳɳ��� | ������ | �ǻ���Ŀ | �������� | Ѫ��Ũ��mg% | ������ | ���� |
| �� | ��ά����ԭ��Fibrinogen) | �� | 340,000���ˣ�ţ�� | 3��2 | 3-4 | 200-400 | ��ά���� | �γ����� |
| �� | ��Ѫøԭ��Prothrombin) | �� | 68,700���ˣ�
72,000��ţ�� |
1 | 8.2���ˣ�
10-14��ţ�� |
10-15 | ��Ѫø | ����ø |
| �� | ��֯��Ѫ���أ�Tissue Thromboplastin) | ����֯ϸ�� | 330,00
220,000��ţ�� |
�� | �� | �� | �� | ������ |
| �� | �����ӣ�Calcium Ion) | �� | �� | �� | �� | �� | �� | ������ |
| �� | ǰ�����أ�Proaccelerin) | �� | 290,000-400,000 | ��� | 11-18 | 5-10 | ����Va) | ������ |
| �� | Ѫ����Ѫ��øת�������(Convertin)�ֳ�SPCA�� | �� | 63,000���ˣ� | 1 | 9.1 | 0.4-0.7 | ��a | ����ø |
| �� | �����ֹw�Բ���(Antihemotpilic Globulin��дAHG�� | ��Ϊ�� | 1,100,000���ˣ�ţ�� | �� | 6���ˣ�
9��ţ�� |
15-20 | ��a | ������ |
| �� | Ѫ����Ѫ���سɷ�(Plasma Thromboplastin��дPTC)����������Ѫ�Ѳ����� | �� | 55,400���ˣ�ţ�� | 1 | 26 | 3-5 | ��a | ����ø |
| �� | Stuart-Prower���� | �� | 55,000���ˣ�ţ�� | 1 | 10 | 5-10 | ��a | ����ø |
| �� | Ѫ����Ѫ����ǰ��(Plasma Thro mboplastin Antecedent��дPTA)����������Ѫ�Ѳ����� | �Σ���״��Ƥϵͳ�� | 160,000���ˣ�ţ�� | 2 | 12 | 0.5-0.9 | ��a | ����ø |
| �� | ������(Hageman���ӣ� | ��״��Ƥϵͳ�� | 90,00��ţ��
82,000���ˣ� |
3 | 15 | 0.1-0.5 | ��a | ����ø |
| X�� | ��ά�����ȶ�����(Fibrin Stabilizing Factor��дFSK�� | Ѫˮ�壿�Σ� | 320,000��Ѫ����
146,000-165,000 ������ѪС�壩 |
5��Ѫ���� | 1-2 | ��a | �γ��ż� | �� |
| �� | ǰ�����ͷ�ø(Prekallidrein) | �� | 80,000 | 1 | 10 | 1-2 | �����ͷ�ø | ����ø |
| �� | �߷���������ԭ(High Molecular Weight Kininogen HMWK) | �� | 110,000-15,000 | 1 | ? | 7 | ������ | ������ |
�����������ֺ��a��ʾ�����˼��activated),���Ӣ�ʵ���Ͼ��Ǣ�a��
����(һ)��Դ��;��
������Դ��;���漰������Ѫ���ӻ���ɷ�Ϊ������
����1.�Ӵ���������Ӣ���Ҳ��Hagemann�����ӵļ������á��˵������ڽӴ����ɸ���ı��棬�粣���������ڽӴ�����ԭ����ʱ����������ı䣬��������Ӣ�aΪһ����ø���ܽ������ͷ�øԭת��Ϊ�����ͷ�ø���ֿɻ���Ӣ����γ�һ����������ͬʱ���Ӣ�a���ɼ�����һ�����Ӣ�������ת��Ϊ��a�����⣬�ڢ����ӻ�л��и߷���������ԭ(high molecular weight kininogen,HMWK)�IJ���(ͼ10��2)��

ͼ10��2����Դ����Ѫ�ĽӴ����
����2.��֬������Ӧ�Σ���Ģ�����a���������Ӣ�����Ca�����Ĵ�����ˮ�����Ӣ�������a�����Ӣ�a��ø���ԣ�����ʹ����X�Ļ��Ӧ�ٶ����1000�����������X(��Xa)����Ѫø���м������Ӣ��͢������á��������Xa��Va��Ca�����������֬�������γ���Ѫøԭ�������֬��������ѪС���ṩ�ĸ���˿������֬��֬���ף�����Ѫ���Ӻ�Ca�����н�ǿ�������������Ӣ��Ļ����Ѫøԭ�����������һϵ�з�Ӧ�����������Ͻ��У��ʳ���֬������Ӧ�Ρ�(ͼ10��3)
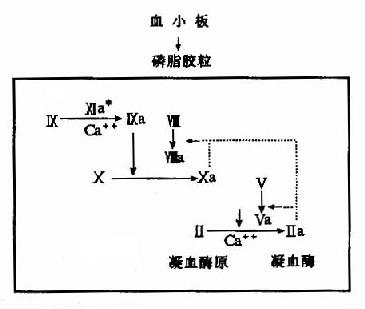
ͼ10��3����Դ����Ѫ����֬������Ӧ��
��(*��Ҫ����Դ����Ѫ�����������Ӣ�a���ʹ���ӻ�����ߡ���ʾ��������·)
����(��)��Դ��;��
��֯���˺��ͷ����Ӣ�(��֯��Ѫ����)������һ��֬���ף����ԡ��Ρ�̥�̵���֯�к�����ḻ��������֬��������ѪС�����ṩ����֬�������ܰ�Ѫ������Ѫ���Ӣ���Xͨ��Ca�����Ŷ������������ϡ����Ӣ����ɢ�a����Ѫø�����ɱ�Xa�����a�ɼ�������X����Xa������֯��Ѫ���صĵ��ײ��ֿ�ʹ�˷�Ӧ����16��000����δ������Ӣ�Ҳ���д����ã������Т�a��2%(ͼ10��4)��
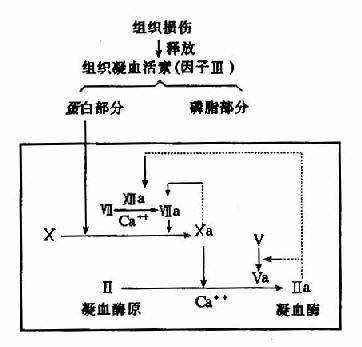
ͼ10-4����Դ����Ѫ����Ѫøԭ����������ɼ���Ѫø����(����ʾ��������·)
����������Ѫøԭ�ļ���
��Ѫøԭ(��prothrombin)�Ǻ�582������л���øԭ��������Xa��Arg��Thr��Arg��Ile���п����г�N����274��������л�������308��������л��ֳ�A��B������������һ���������������Ϊ��Ѫø(thrombin)��(ͼ10��5)����Va��ø���ԣ�����ʹXa�Ļ�����ǿ350����������Ѫø�����ɡ���֬������ø(Xa)�͵���(��Ѫøԭ)֮���Ca������Ϊ������������Ѫøԭ������N��δ�˺���10���ê��Ȼ��Ȱ���л������ڵ��Ȼ�����Ca�����γɸ����塣��һ���棬Ca�����ֿ�����֬���������ϣ�����ʹXa��Va����Ѫøԭ�Ӵ���һ������Xa����Ѫøԭˮ��Ϊ��Ѫø(ͼ10��6)��

ͼ10��5������Xa������Ѫøԭʾ��ͼ
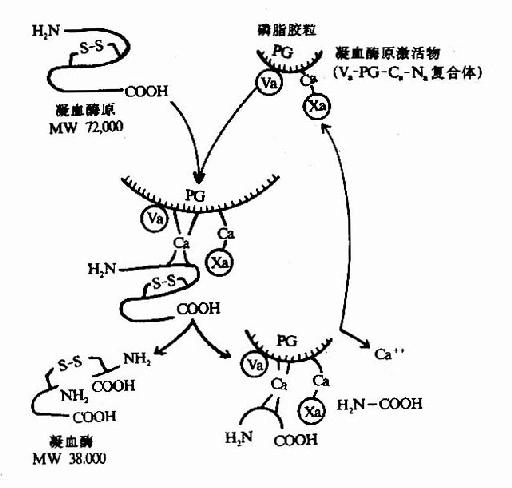
ͼ10��6����Ѫø������
������Ѫøԭ�����Ӣ������������ɸκϳɣ��ϳɹ�������Ҫά��K��Ϊ�����ӡ�ȱ��Vitk�������쳣��Ѫøԭ��ֻ���������Ե�1?%���о�����Vitk������Ѫøԭ�ê��Ȼ��Ȱ�������ɡ�Vitk�����Ȼ����Ļ���Ϊ��������Vitk��ø�Ĵ��¶�ȥ�ê�C�ϵ�һ�����ӣ�ʹ�ã�C�������ӣ�����CO2��ϡ�2��3������Vitk����ø���±������ỹԭ���ظ����ã����Vitk�ڴ��Ȼ���Ӧ����ø�����á�(ͼ10��7)

ͼ10��7��ά����K�ڹȰ���л�r���Ȼ���Ӧ�е�����(ά����Kѭ��)
�����ġ���ά����ԭת��Ϊ��ά����
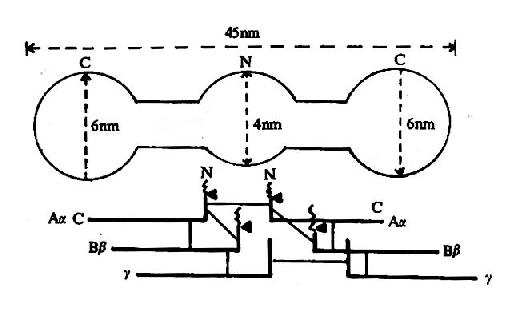
ͼ10��8����ά�ص���ԭ����ʾ��ͼ��
�ϰ�Ϊ�羵�µķ�����״���°�ʾ6������������
һΪ˫�������Ϊ��Ѫø���õ�

ͼ10��9����ά�ص�������������
����ѪҺ���̵�ʵ������ά�������������ɣ�����Ѫ������ά����ԭ(fibrinogen)����Ѫø�����½���Ϊ��ά���ײ��ۺϳɲ����Ե���״�ṹ��
������ά����ԭ���������Ԧ��������£������ã�����ɣ�ÿ3������(�����¡���)�ʺϳ���״���γ�������״��������Nĩ���ж����ʹ̬�����ӵõ��ȶ���������������N���˷ֱ���һ��16����14���������С�ģ���Ϊ��ά��A��B����ˣ���ά����ԭ��дΪ(A��B�¦�)2(ͼ10-8)��
������Ѫø�ı���Ϊһ�ֵ���ˮ��ø����������������A����B�����ϵľ������ļ����г�A��B��ά�ġ�����ά��A��B��Ϊ�����ģ����϶ฺ��ɡ����ڵ���ų������谭��ά����ԭ֮��ۺϡ��г���ά��A��Bת��Ϊ��ά�����Լ�С��ͬʱ��¶�˻������λ�㣬�������Զ��ۺϣ���ά����λͨ���߿��ߡ��˿��˵ľۺ��γɾۺ��������ֶ���岻�ȶ�����Ϊ������(soft clot)������ͨ������X��a�����ý���ι̵���������X��aΪת��ø���ܴ�һ������ĹȰ���л��Ħã��Ȼ�����һ�����������л��İ���֮���γɹ��۽�ϣ�����ͳ�NH3(ͼ10��9��10)����ˣ�����X��a��Ϊ��ά�����ȶ�����(fibrin stabilizingfactor,FSF)������X�������ѪС�弰Ѫ���У�����Ѫø�г������Ķκ���ΪX��a��
�����ɴ˲������ȶ���ά�������������鲻ͬ��������5M���弰1%��������Һ�в��ܽ⡣��ѪС���Ѫ˨�������������£�����״�ṹ�����������˿ڱ�Ե�˴˿����������˿ڱպϡ�����άϸ���ı������һ��������ά���ĵ����ʣ���ճ�����ף����ɢ�a������ά���������������ά���̶����������ԣ����Ӣ�a��ֱ�Ӳ����˿ڵ����ϡ�
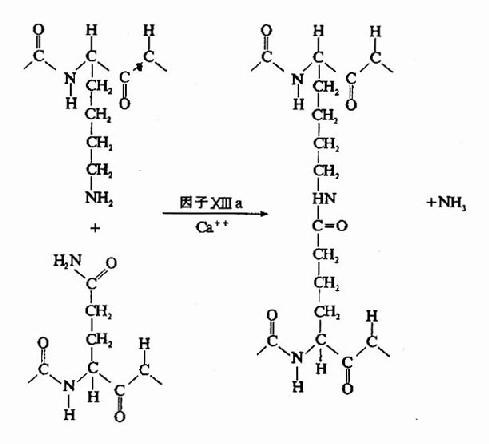
ͼ10��10������X��a���û���
�����ܽ�������Ѫ���̿ɹ��ɳ������ص㣺
����1.��Ѫ���ӵĻ������Ϊ�����ʵ�����ˮ�⣬��������Ѫ���˱������ǵ���ø������ø���õĵ����Щ����Ϊ����ø����Ѫ����(��������������)�İ�����˳������ƣ�������˿���ᵰ��øͬԴ���������ĵ�˿����л������ļ���ˮ�⡣C����Լ250��������л�ͬԴ�Ժܸߣ��Ǿ��д����ԵĽṹ��N���˵İ��������б仯�ϴ�������Ѫ�������õ����רһ�ԡ����Ǵ��ķ�Ӧ��Ca��������֬�μӡ�
ͼ10��11��ѪҺ���̵��ٲ�ЧӦ
����2.��֬����(��Դ��;����ѪС�壬��Դ��;������֯��Ѫ�����ṩ)ʹ���Ӧ�ڽ���������У������߷�Ӧ�ٶȣ���Ca�������������ڴٽ�ø�͵�������֬����Ľ�ϡ�
����3.��Ѫ���ӻ���ٲ�ЧӦ(cascade)ʹѪҺ���̾��и�Ч�ʺ;��ܵ��ص���������ͼ10��11��ʾ��
����4.ά����K���ڡ���Դ����Ѫ�о�����Ҫ���á�
����5.��Ѫ�����е�������ʹ��Ӧ���ϼ��٣����ղ�����ά�����п���Ѫ���á���������Ѫ�뿹��Ѫ��������ϵ�ġ�
�����塢��Ѫ���õĵ���
����������������Ѫ������һ�������Ŵ���ٲ�ЧӦ����֮���������ã��ɰ�������ɵ�ø���Լ�����ǿ�������в������������ǿ106������˸ߵļ����ٶȻ�Ի��幹��Σ�գ�����˵���˹���һ������������ѪҺ�ͻ��������������⣬Ѫ��������ļ���������Ѫ˨�����ؼ�������ˣ������ڵ���Ѫ���ñ��뱣���ʶȡ�ʵ���ϣ�Ѫ����Ѫ����Ƥ�ȴ������Ŷ��ֿ������ʣ���Ѫ���������ɵ���ά����(����Ѫø��)��ǿ��������Ѫø�����á�Ѫ���п���Ѫ����(antithrombin����Ѫø��)��һ�ַ�����Լ58��000���ǵ��ף�������е���ø���õ���Ѫ����(��a��IXa��Xa����a����a)��1��1���ӱȽ���γɸ�����Ӷ����ø�Ļ������ġ�����(heparin)�ܼ��ٸ�������γɣ�ʹ����Ѫø�Ļ���������ٱ����������ɷʴ�ϸ�����ȼ�����ϸ�������ĸ߷�������ճ���ǣ���һ����Ҫ����Ѫ���ʡ������������⣬�о�������ѪС���ճ�������ۣ��Ӷ�Ӱ��ѪС����֬���ͷŵ����á�������Ϊ�������ѹ㷺Ӧ�����ٴ���
ͼ10��12�����ֿ������ʽṹ
����Ѫ���л�������һ�ֿ���Ѫ�ĵ����ʗC���ס���Gla�л����Ƿ�����Լ62��000���ǵ��ף���øԭ��ʽ���ڣ�����Ѫø�������ˮ���a����a���Ӷ����ӿ���Ѫ���ܡ�������ȱ��C�����ߣ�������Ӥ���ڼ����ڹ㷺��Ѫ˨��
��������Ȼ������Ѫ���еĿ��������⣬�ٴ��ϳ���һЩ�˹�������������κ��������Σ����ǵ�������ͨ������ȥ��Ca���������˫�㶹�����������Vitk�������ӿ���Ѫ����(ͼ10��12)��
����������ά�����ܽ�
����ѪҺ��������������ά���ɱ�Ѫ������ά������øϵͳ�����ܽ⣬���ڷ�ֹѪ˨�γɺͱ���Ѫ��ͨ��������Ҫ���塣�����˵�һЩ����Һ(����֭����Һ����Һ���ӹ��������������Һ��)�о�������ά������øԭ(plasminogen)�����������ά�����ܽ���̣���ʱ������ڹܵ��ڵ���ά���ף��Ա��ַ��ڹܵ���ͨ�����¾�ѪҺ�������ԡ�
����(һ)��ά�����ܽ����
������ά�����ܽ�(fibrinolysis)���̿ɷ�Ϊ���࣬����ά����øԭ�������ά�����ܽ⡣
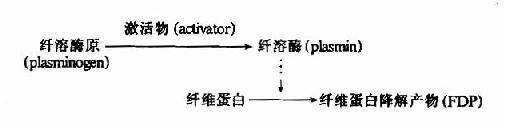
����1.��ά����øԭ�ļ���
����(1)��ά����øԭ�����
����ѪҺ����ά����ø(plasmin���������ø)������øԭ��ʽ���ڣ�ֻ�������ܼ�����������ת��Ϊ����ø�ž��л��ԡ����ܼ�����ɷ�Ϊ��֯�������ѪҺ�����������ࡣ
������֯����������������֯ϸ����ø���У����ӹ���ǰ���١���״�١��Ρ����Ⱥ����϶ࡣ�����о�����õ������е���ø(urokinase)�������������������Һ�ж�����������С��Ѫ˨���ܽ������Ҫ���á�
����ѪҺ��������Ҫ���Ծ�����������Ѫ����Ƥϸ�������ܵ�ijЩ�̼�(������˶����������š����ˡ��ݿ˵�)ʱ���ɴ�ʹ��Ƥϸ���ϳ����ಢ�ͷ���Ѫ����֪ѪС���ͷ�5�HT��Ѫ����Ƥϸ���ͷ����ܼ����������Ҫ���塣
�������⣬����Ѫ���������ȡ��һ�ֵ����ʳ�Ϊ����ø(streptokinase)����������øԭ�γɸ�������߾������ܼ���������ʡ���ø������øĿǰ�ѹ㷺Ӧ�����ٴ���˨���ơ�
����(2)����øԭ�ļ���
��������øԭΪ������86��000�ĵ��ס����ܼ������Ϊ����ˮ��ø����ˮ������øԭʹ֮��������Arg��Val���ж϶����������ø������ø������ɻ����øԭ��ͬʱ����ˮ����ά����ԭ�����Ӣ�������IX�͢��ȣ��Ӷ�������Ѫ��
����2.��ά�����ܽ�
������ά�����ܽ�����Ƿֲ����еģ����ȱ�����øˮ���ͳ�A��B��CС���Ӷ��ģ�����XƬ���Ա����������ԡ�XƬ�Ͻ�һ��ˮ��ΪƬ��D��Y��Y��ˮ��ΪD��EƬ�ϡ�(ͼ10��13)���շֽ����ΪA��B��C��D��E����Ƭ�Ρ���ЩƬ��ͳ��Ϊ��ά���������(fibrin degradation product,FDP)��FDP�����������ǣ�Ƭ��X��Y������ά������ۺϣ����ƶ��������ɣ�Ƭ��D��ֱ��������ά������ľۺϣ�Ƭ��Y��E��ɾ���������Ѫø�����ң���FDP�ɸ���ѪС���ճ�����ۼ����ɼ�FDP�ڿ���������Ҫ���á�
����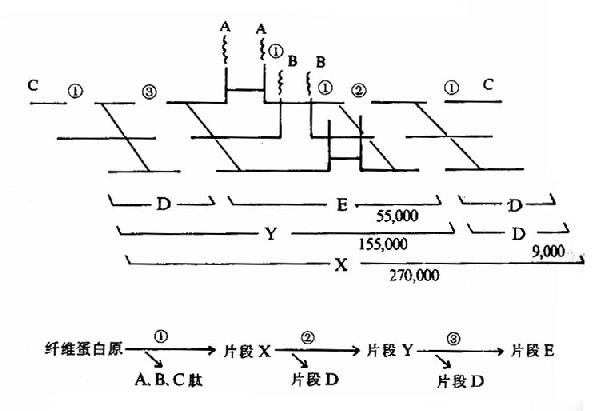
ͼ10��13������ø����ά����(ԭ)�Ľ���(Ƭ���µ�����Ϊ������)
����(��)����������
����������֯����Һ�㷺��������������������ÿɷ�Ϊ������øԭ��������������ø������ֳƿ�����ø(antiplasmin)��
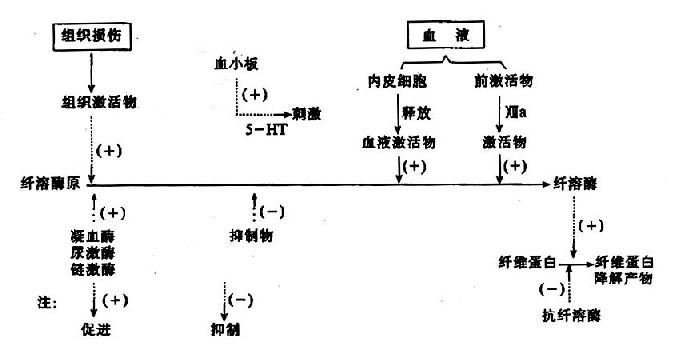
ͼ10��14����ά�����ܽ����

ͼ10��15����Ѫȫ����ʾ��ͼ(��ʾת����ͷţ���ʾ�����ã�
���Ӵ�����⣬����Ca�������룬ͼ��δ��ʾ)
��������ѪҺ�п�����ø����������ø���Ե�20?0�����������������£�����ø���Է������á�������ø�����֣��������õĿ���άø������1���ȵ���ø��������Ϊ45��000����������ø�γ��ι̵ĸ�����ڿ����ÿ�����ø������2���ף�������80��000��������ø�ľ������Ƽ���
����ѪС���������������ǿ������õĿ�����ø����Ѫ˨�γ����ڣ�ѪС�������������ô������ܼ������ã���Ѫ˨�����Ժ�����ѪС����5�HT���ͷţ���Ѫ����Ƥ�ͷ�ѪҺ���������࣬��������ά�����ܽ⣬��ֹѪ˨��������������Ѫ��ѭ����
������˿ɼ�������ѪҺ���̡������������뿹����������ơ��Э������ͬά��ѪҺϵͳ�������������ܣ������ϵ�ɼ�ͼ10��14��17��
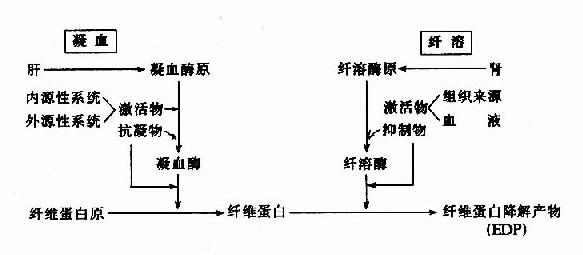
����ͼ10��16����Ѫ�����ܵıȽ�

ͼ10��17����Ѫ�����ܵ����ϵ