第三节 气体在血液中的运输
从肺泡扩散入血液的O2必须通过血液循环运送到各组织,从组织散入血液的CO2的也必须由血液循环运送到肺泡。下述O2和CO2在血液中运输的机制。
一、氧和二氧化碳在血液中存在的形式
O2和CO2的都以两种形式存在于血液:物理溶解的和化学结合的。
气体在溶液中溶解的量与分压和溶解度成正比,和温度成反比。温度38℃时,1个大气压(760Hg,101.08kPa)的 O2和 CO2和在100ml血液中溶解的量分别是2.36ml和48ml。按此计算,静脉血 PCO2和为6.12kPa(46mmHg),则每100ml血液含溶解的CO2为(48×6.12)/101.08=2.9ml;动脉血PO2为13.3kPa(100mmHg),每100ml血液含溶解的O2为(2.36×13.3)/101.08=0.31ml。可是,血液中实际的O2和O2为CO2含量比这数字大得多(表5-4),以溶解形式存在的O2、CO2比例极少,显然单靠溶解形式来运输O2、CO2不能适应机体代谢的需要。例如,安静状态下人体耗O2量约为250ml/min,如只靠物理溶解的O2来提供,则需大大提高心输出量或提高肺泡内的PO2,这对机体极其不利,所幸在进化过程中形成了O2、CO2为极为有效地化学结合的运输形式,大大减轻了对心脏和呼吸器官的苛求。
表5-4血液O2和CO2的含量(ml/100ml 血液)
| 物理溶解的 | 动脉血 化学结合的 |
合计 | 物理溶解的 | 混合静脉血 化学结合的 |
合计 | |
| O2 | 0.31 | 20.0 | 20.31 | 0.11 | 15.2 | 15.31 |
| CO2 | 2.53 | 46.4 | 48.93 | 2.91 | 50.0 | 52.91 |
虽然溶解形式的O2、CO2很少,但也很重要。因为在肺或组织进行气体交换时,进入血液的O2、CO2都是先溶解,提高分压,再出现化学结合;O2、CO2从血液释放时,也是溶解的先逸出,分压下降,结合的再分离出现补充所失去的溶解的气体。溶解的和化学结合的两者之间处于动态平衡。
二、氧的运输
血液中的O2以溶解的和结合的两种形式存在。溶解的量极少,仅占血液总O2含量的约1.5%,结合的占 98.5%左右。O2的结合形式是氧合血红蛋白(HbO2)。血红蛋白(hemoglobin,Hb)是红细胞内的色蛋白,它的分子结构特征使之成为极好的运O2工具。Hb还参与CO2的运输,所以在血液气体运输方面Hb占极为重要的地位。
(一)Hb分子结构简介
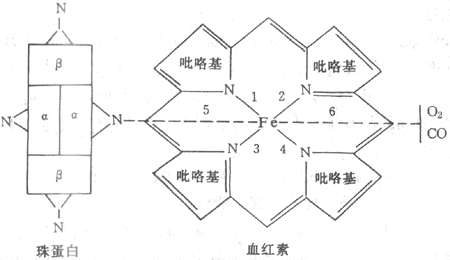
每1Hb分子由1个珠蛋白和4个血红素(又称亚铁原卟啉)组成(图5-12)。每个血红素又由4个吡咯基组成一个环,中心为一铁原子。每个珠蛋白有4条多肽链,每条多肽链与1个血红至少连接构成Hb的单体或亚单位。Hb是由4个单体构成的四聚体。不同Hb分子的珠蛋白的多肽链的组成不同。成年人Hb(HbA)的多肽链是2条α链和2条β链,为α2β2结构。胎儿Hb(HbF)是2条α链和2条γ链,为α2γ2结构。出生后不久HbF即为HbFA所取代。多肽链中氨基酸的排列顺序已经清楚。每条α链含141个氨基酸残基,每条β链含146个氨在酸残基。血红素的Fe2+均连接在多肽链的组氨基酸残基上,这个组氨酸残基若被其它氨基酸取代,或其邻近的氨基酸有所改变,都会影响Hb的功能。可见蛋白质结构和功能密切相关。
Hb的4个单位之间和亚单位内部由盐键连接。Hb与O2的结合或解离将影响盐键的形成或断裂,使Hb四级结构的构型发生改变,Hb与O2的亲和力也随之而变,这是Hb氧离曲线呈S形和波尔效应的基础(见下文)。
图5-12 血红蛋白组成示意图
(二)Hb与O2结合的特征
血液中的O2主要以氧合Hb(HbO2)形式运输。O2与Hb的结合有以下一些重要特征:
1.反应快、可逆、不需酶的催化、受PO2的影响。当血液流经PO2高的肺部时,Hb与 O2结合,形成HbO2;当血液流经PO2低的组织时,HbO2迅速解离,释放O2,成为去氧Hb:

2.Fe2+与O2结合后仍是二价铁,所以该反应是氧合(oxygenation),不是氧化(oxidation)。
3.1分子Hb可以结合4分子O2。Hb分子量是64000-67000道尔顿(d),所以1gHb可以结合1.34-1.39mlO2,视Hb纯度而异。100ml血液中,Hb所能结合的最大O2量称为Hb的氧容量。此值受Hb浓度的影响;而实际结合的O2量称为Hb的氧含量,其值可受PO2的影响。Hb氧含量和氧容量的百分比为Hb氧饱和度。例如,Hb浓度在15g/100ml血液时,Hb的氧容量=15×1.34=Hb 20.1ml/100ml血液,如Hb的氧含量是20.1ml,则Hb氧饱和度是100%。如果Hb氧含量实际是15ml,则Hb氧饱和度=15/20×100%=75%。通常情况下,溶解的O2极少,故可忽略不计,因此,Hb氧容量,Hb氧含量和Hb氧饱和度可分别视为血氧容量(osygen capacity)、血氧含量(oxygen content)和血氧饱和度(oxygen saturatino)。HbO2呈鲜红色,去氧Hb呈紫蓝色,当体表表浅毛细血管床血液中去氧Hb 含量达5g/100ml血液以上时,皮肤、粘膜呈浅蓝色,称为紫绀。
4.Hb与O2的结合或解离曲线呈S形,与Hb的变构效应有关。当前认为Hb有两种构型:去氧Hb为紧密型(tense form,T型),氧合Hb为疏松型(relaxed form,R型)。当O2与Hb的Fe2+结合后,盐键逐步断裂,Hb 分子逐步由T型变为R型,对O2的亲和力逐步增加,R型的O2亲和力为T型的数百倍。也就是说,Hb 的4个亚单位无论在结合O2或释放O2时,彼此间有协同效应,即1个亚单位与O2结合后,由于变构效应的结果,其它亚单位更易与O2结合;反之,当HbO2的1个亚单位释出O2后,其它亚单位更易释放O2。因此,Hb氧离曲线呈S形。
(三)氧离曲线
氧离曲线(oxygen dissociation curve)或氧合血红蛋白解离曲线是表示PO2与Hb 氧结合量或Hb氧饱和度关系的曲线(图5-13)。该曲线既表示不同PO2时,O2与Hb 的结合情况。上面已经提到的曲线呈S形,是Hb变构效应所致。同时曲线的S形还有重要的生理意义,下面分析氧离曲线各段的特点及其功能意义。
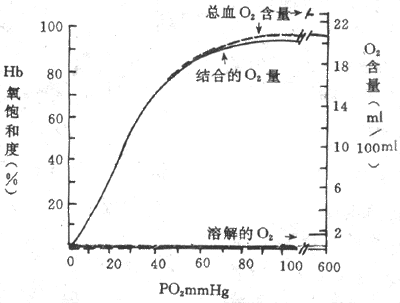
图5-13 氧离曲线
(实线,在Ph7.4,PCO2 40mmHg,温度37℃时测定的)
同时示溶解的O2和在Hb浓度为15g/100ml血液时的总血O2含量(1mmHg=0.133kPa)
1.氧离曲线的上段 相当于PO27.98-13.3kPa(60-100mmHg),即PO2较高的水平,可以认为是Hb与O2结合的部分。这段曲线较平坦,表明PO2的变化对Hb氧饱和度影响不大。例如PO2为13.3kPa(100mmHg)时(相当于动脉血PO2),Hb氧饱和度为97.4%,血O2含量约为19.4ml%;如将吸入气PO2提高到19.95kPa(150mmHg),Hb氧饱和度为100%,只增加了2.6% ,这就解释了为何VA/Q不匹配时,肺泡通气量的增加几乎无助于O2的摄取;反之,如使PO2下降到9.31kPa(70mmHg),Hb氧饱和度为94%,也不过只降低了3.4%。因此,即使吸入气或肺泡气PO2有所下降,如在高原、高空或某些呼吸系统疾病时,但只要PO2不低于7.98kPa(60mmHg),Hb氧饱和度仍能保持在90%以上,血液仍可携带足够量的O2,不致发生明显的低血氧症。
2.氧离曲线的中段 该段曲线较陡,相当于PO25.32-7.98kPa(40-60mmHg),是HbO2释放O2的部分。PO25.32kPa(40mmHg),相当于混合静脉血的PO2,此时Hb氧饱和度约为75%,血O2含量约14.4ml%,也即是每100ml血液流过组织时释放了5mlO2。血液流经组织液时释放出的O2容积所占动脉血O2含量的百分数称为O2的利用系数,安静时为25%左右。以心输出量5L计算,安静状态下人体每分耗O2量约为250ml。
3.氧离曲线的下段 相当于PO22-5,32kPa(15-40mmHg),也是H bO2与O2解离的部分,是曲线坡度最陡的一段,意即PO2稍降,HbO2就可大大下降。在组织活动加强时,PO2可降至2kPa(15mmHg),HbO2进一步解离,Hb氧饱和度降至更低的水平,血氧含量仅约4.4ml%,这样每100ml血液能供给组织15mlO2,O2的利用系数提高到75%,是安静时的3倍。可见该段曲线代表O2贮备。
(四)影响氧离曲线的因素
Hb与O2的结合和解离可受多种因素影响,使氧离曲线的位置偏移,亦即使Hb对O2的亲和力发生变化。通常用P50表示Hb对O2的亲和力。P50是使Hb氧饱和度达50%时的PO2,正常为3.52 kPa(26.5mmHg)。P50增大,表明Hb对O2的亲和力降低,需更高的PO2才能达到50%的Hb氧饱和度,曲线右移;P50降低,指示Hb对O2的亲和力增加,达50%Hb氧饱和度所需的PO2降低,曲线左移。影响Hb与O2亲和力或P50的因素有血液的Ph、PCO2、温度和有机磷化物(图5-14)。
1.Hb与PCO2的影响pH降低或升PCO2升高,Hb对O2的亲和力降低,P50增大,曲线右移; pH升高或PCO2降低,Hb对O2的亲和力增加,P50降低,曲线左移。酸度对Hb氧亲和力的这种影响称为波尔效应(Bohr effect)。波尔效应的机制,与 pH改变时h b构型变化有关。酸度增加时,H+与Hb多肽链某些氨基酸残基的基团结合,促进盐键形成,促使Hb分子构型变为T型,从而降低了对O2的亲和力,曲线右移;酸度降低时,则促使盐键断裂放出H+,Hb变为R型,对O2的亲和力增加,曲线左移。PCO2的影响,一方面是通过PCO2改变时,pH也改变间接效应,一方面也通过CO2与Hb结合而直接影响Hb与O2的亲和力,不过后一效应极小。
波尔效应有重要的生理意义,它既可促进肺毛细血管的氧合,又有利于组织毛细血管血液释放O2。当血液流经肺时,CO2从血液向肺泡扩散,血液PCO2下降,[H+]也降低,均使Hb对O2的亲和力增加,曲线左移,在任一PO2下Hb氧饱和度均增加,血液运O2量增加。当血液流经组织时,CO2从组织扩散进入血液,血液PCO2和[H+]升高,Hb对O2的亲和力降低,曲线右移,促使HbO2解离向组织释放更多的O2。
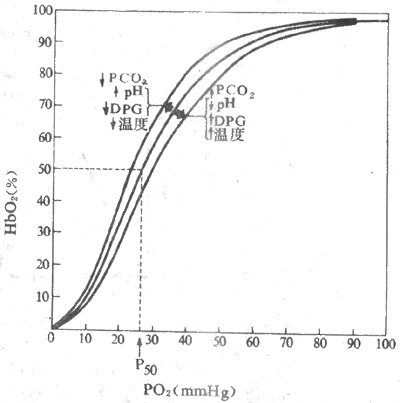
图5-14 影响氧离曲线位置的主要因素(1mmHg=0.133kPa)
2.温度的影响 温度升高,氧离曲线右移,促使O2释放;温度降低,曲线左移,不利于O2的释放。临床低温麻醉手术时应考虑到这一点。温度对氧离曲线的影响,可能与温度影响了H+活度有关。温度升高H+活度增加,降低了Hb对O2的亲和力。当组织代谢活跃是局部组织温度升高,CO2和酸性代谢产物增加,都有利于Hb02解离,活动组织可获得更多的O2以适应其代谢的需要。
3.2,3-二磷酸甘油酸 红细胞中含有很多有机磷化物,特别是2,3-二磷酸甘油酸(2.3-diphospoglyceric acid,2,3-DPG),在调节Hb和O2的亲和力中起重要作用。2,3-DPG浓度升高,Hb对O2亲和力降低,氧离曲线右移:2,3-DPG浓度升降低,Hb对O2的亲和力增加,曲线左移。其机制可能是2,3-DPG与Hbβ链形成盐键,促使Hb变成T型的缘故。此外,2,3-DPG可以提高[H+],由波尔效应来影响Hb对O2的亲和力。
2,3-DPG是红细胞无氧糖酵解的产物。高山缺O2,糖酵解加强,红细胞 2,3-DPG增加,氧离曲线右移,有利于O2的释放,曾认为这可能是能低O2适应的重要机制。可是,这时肺泡PO2也降低,红细胞内过多的2,3-DPG也妨碍了Hb与O2的结合。所以缺O2时,2,3-DPG使氧离曲线右移是否有利,是值得怀疑的。
4.Hb自身性质的影响 除上述因素外,Hb与O2的结合还为其自身性质所影响。Hb的Fe2+氧化成Fe3+,失去运O2能力。胎儿Hb和O2的亲和力大,有助于胎儿血液流经胎盘时从母体摄取O2。异常Hb 也降低运O2功能。CO与Hb结合,占据了O2的结合位点,HbO2下降。CO与Hb的亲和力是O2的250倍,这意味着极低的PCO,CO就可以从HbO2中取代O2,阻断其结合位点。此外,CO还有一极为有害的效应,即当CO与Hb分子中某个血红素结合后,将增加其余3个血红素对O2的亲和力,使氧离曲线左移,妨碍O2的解离。所以CO中毒既妨碍Hb与O2的结合,又妨碍O2的解离,危害极大。
总之,血液Hb的运O2量可受多种因素影响:包括PO2、Hb本身的性质和含量、pH、PCO2、温度、2,3-DPG和CO等,pH降低,PCO2升高,温度升高,2,3-DPG增高,氧离曲线右移;pH升高,PCO2、温度、2,3-DPG降低和CO中毒,曲线左移。
三、二氧化碳的运输
(一)CO2的运输
血液中CO2也 以溶解和化学结合的两种形式运输。化学结合的CO2主要是碳酸氢盐和氨基甲酸血红蛋白。表5-5示血液中各种形式CO2的含量(ml/100ml 血液)、运输量(%)和释出量(%)。溶解的CO2约占总运输量的5%,结合的占95%(碳酸氢盐形式的占88%,氨基甲酸血红蛋白形式占7%)。
从组织扩散入血CO2首先溶解于血浆,一小部分溶解的CO2缓慢地和水结合生成碳酸,碳酸又解离成碳酸氢根和氢离子,H+被血浆缓冲系统缓冲,pH无明显变化。溶解的CO2也与血浆蛋白的游离氨基反应,生成打官司基甲酸血浆蛋白,但形成的量极少,而且动静脉中的含量相同,表明它对CO2的运输不起作用。
在血浆中溶解的CO2绝大部分扩散进入红细胞内,在红细胞内主要以下述结合形式存在:
表5-5 血液中各种形式CO2的含量(ml/100ml血液)、运输量(%)和释出量(%)
| 动脉血 | 静脉血 | 差值 | 释出题 | |||
| 含量 | 运输量 | 含量 | 运输量(动、静永血间) | |||
| CO2总量 | 48.5 | 100 | 52,5 | 100 | 4.0 | 100 |
| 溶解的CO2 | 2.5 | 5.15 | 2.8 | 5.33 | 0.3 | 7.5 |
| HCO3 形式的CO2 | 43.0 | 88.66 | 46.0 | 87.62 | 3.0 | 75 |
| 氨基甲酸血红
蛋白的CO2 |
3.0 | 6.19 | 3.7 | 7.05 | 0.7 | 17.5 |
运输量(%)是指各种形式的CO2含量/CO2总含量×100%
释放量(%)是指各种形式的CO2在肺释放量/CO2总释放量×100%
1.碳酸氢盐 从组织扩散进入血液的大部分CO2,在红细胞内与水反应生成碳酸,碳酸又解离成碳酸氢根和氢离子,反应极为迅速,可逆(图5-15)。这是因为红细胞内含有较高浓度的碳酸酐酶,在其催化下,使反应加速5000倍,不到1s即达平衡。在此反应过程中红细胞内碳酸氢根浓度不断增加,碳酸氢根便顺浓度梯度红细胞膜扩散进入血浆。红细胞负离子的减少应伴有同等数量的正离子的向外扩散,才能维持电平衡。可是红细胞膜不允许正离子自由通过,小的负离子可以通过,于是,氯离子便由血浆扩散进入红细胞,这一现象称为氯离子转移(chloride shift)。在红细胞膜上有特异的HCO3—CI-载体,运载这两类离子跨膜交换。这样,碳酸氢根便不会在红细胞内堆积,有利于反应向右进行和CO2的运输。在红细胞内,碳酸氢根与K+结合,在血浆中则与Na+结合成碳酸氢盐。上述反应中产生的H+,大部分和Hb结合,Hb 是强有力的缓冲剂。
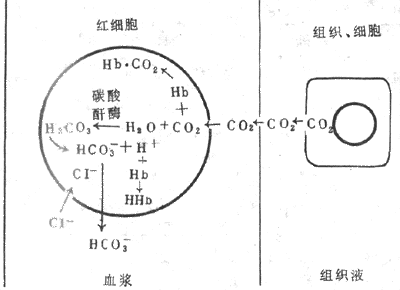
图5-15 CO2在血液中的运输示意图
在肺部,反应向相反方向(左)进行。因为肺泡气PCO2比静脉血的低,血浆中溶解的CO2首先扩散入肺泡,红细胞内的HCO3+H+生成H2CO3,碳酸酐酶又催化H2CO3分解成CO2和H2O,CO2又从红细胞扩散入血浆,而血浆中的HCO3便进入红细胞以补充消耗的HCO3,CI-则出红细胞。这样以HCO3形式运输的CO2,在肺部又转变成CO2释出。
2.氨基甲酸血红蛋白 一部分CO2与Hb的氨基结合生成氨基甲酸血红蛋白(carbaminohemoglobin),这一反应无需酶的催化、迅速、可逆,主要调节因素是氧合作用。
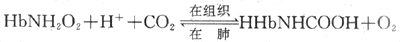
HbO2与CO2结合形成HbNHCOOH的能力比去氧Hb的小。在组织里,解离释出O2,部分HbO2变成去氧Hb,与CO2结合生成HbNHCOOH。此外,去氧Hb 酸性较HbO2弱,去氧Hb和H+结合,也促进反应向右侧进行,并缓冲了pH的变化。在肺的HbO2生成增多,促使HHbNHCOOH解离释放CO2和H+,反应向左进行。氧合作用的调节有重要意义,从表5-5可以看出,虽然以氨基甲酸血红蛋白形式运输的CO2仅占总运输量的7%,但在肺排出的CO2中却有17.5%是从氨基甲酸血红蛋白释放出来的。
(二)CO2解离曲线
CO2解离曲线(carbon dioxide dissociation curve)是表示血液中CO2含量与PCO2关系的曲线(图5-16)。与氧离曲线不同,血液CO2含量随PCO2上升而增加,几乎成线性关系而不是s 形,而且没有饱和点。因此,CO2解离曲线的纵坐标不用饱和度而用浓度来表示。
图5-16的A点是静脉血PO25.32kPa(40mmHg),PCO26kPa(45mmHg)时的CO2含量,约为52ml%;B点是动脉血PO213.3kPa(100mmHg),PCO25.32kPa(40mmHg)时的CO2含量,约为48ml%,血液流经肺时通常释出CO24ml/100ml血液。
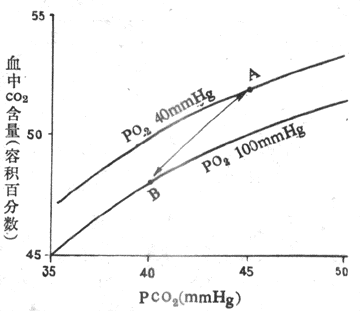
图5-16 CO2解离曲线
A:静脉血 B:动脉血 (1mmHg=0.133kPa)
(三)氧与Hb的CO2运输的影响
O2与Hb结合将促使CO2释放,这一效应称作何尔登效应( Haldane effect)。从图5-16可以看出,在相同PCO2下,动脉血(HbO2)携带的CO2比静脉血少。这主要是因为HbO2酸性较强,而去氧Hb酸性较弱的缘故。所以去氧Hb易和 CO2结合生成 HbNHCOOh ,也易于和H+结合,使H2CO2 解离过程中产生的 H+被及时移去,有利于反应向右进行,提高了血液运输CO2的量。于是,在组织中,由于HbO2释出O2而成去氧Hb,经何尔登效应促使血液摄取并结合CO2;在肺,则因Hb与 O2结合,促使CO2释放。可见O2和CO2的运输不是孤立进行的,而是相互影响的。CO2通过波尔效效影响O2的结合和释放,O2又通过何尔登效应影响CO2的结合和释放。两者都与Hb的理化特性有关