第五节 器官循环
体内每一器官的血流量取决于主动脉压和中心静脉压之间的压力差,又取决于该器官阻力血管的舒缩状态。由于各器官的结构和功能各不相同,器官内部的血管分布又各有特征,因此其血流量的调节除服从前已述的一般规律外,还有其本身的特点。本节叙过心、肺、脑几个主要器官的血液循环特征。关于肾的血液循环特征,将在第八章叙述。
一、冠脉循环
(一)冠脉循环的解剖特点
心肌的血液供应来自左、右冠状动脉。冠状动脉的主干行走于心脏的表面,其小分支以垂直于心脏表面的方向穿入心肌,并在心内膜下层分支成网。这种分支方式使冠脉血管容易在心肌收缩进受到压迫。左、右冠状动脉及其分支的走向可有多种变异。在多数人中,左冠状动脉主要供左心室的前部,右冠状动脉主要供应左心室的后部和右心室。左冠状动脉的血液流经毛细血管和静脉后,主要经由冠状窦回流入右心房,而右冠动脉的血液则主要经较细的心前静脉直接回流入右心室。另外还有一小部分冠脉血液可通过心最小静脉直接流入左、右心房和心室腔内。
心肌的毛细血管网分布极为丰富。毛细血管数和心肌纤维数的比例为1:1。在心肌横截面上,每平方毫米面积内约有2500-3000根毛细血管。因此心肌和冠脉血液之间的物质交换可能很快地进行。冠状动脉之间有侧支互相吻合。在人类,这种吻合支戊内膜下较多。正常心脏的冠脉侧较细小,血流量很少。因此当冠状动脉突然阻塞时,不易很快建立侧支循环,常可导致心肌梗塞。但如果冠状动脉阻塞是缓慢形成的,则侧支可逐渐扩张,并可建立新的侧支循环,起代偿作用。
(二)冠脉血流的特点
在安静状态下,人冠脉血流量为每百克心肌每分钟60-80ml。中等体重的人,总的冠脉血流量为225ml/min,占心输出量的4%-5%。冠脉血流量的多少主要取决于心肌的活动,故左心室单位克重心肌组织的血流量大于右心室。当心肌活动加强,冠脉达到最大舒张状态时,冠脉血流量可增加到每百克心肌每分钟300-400ml。
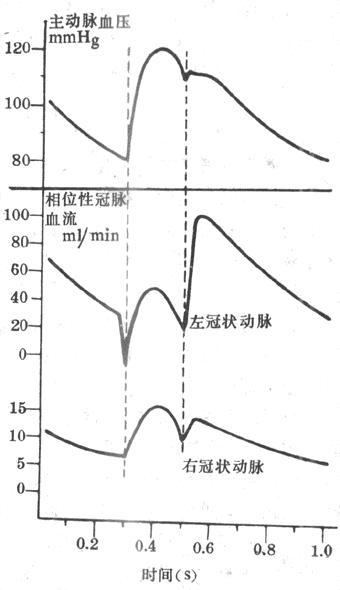
由于肮脏血管的大部分分支深埋于心肌内,心脏在每次收缩时对埋于其内的血管产生压迫,从而影响冠脉血流。图4-29示狗的左、右冠状动脉血流在一个心动周期中的变化。在左心室等容收缩期,由于心肌收缩的强烈压迫,左冠状动脉血流急剧减少,甚至发生倒流。在左心室射血期,主动脉压升高,冠状动脉血压也随着升主,冠脉血流量增加。到慢速射血期,冠脉血流量又有下降。心肌舒张时,对冠脉血管的压迫解除,故冠脉血流的阻力显著减小,血流量增加。在等容舒张期,冠脉血流量突然增加,在舒张期的早期达到最高峰,然后逐渐回降,在左心室深层,心肌收缩对冠血流的影响更为明显。左心房收缩时对冠脉血流也可产生一定的影响,但并不显著。一般说来,左心室在收缩期血流量大约只有舒张期的20%-30%。当心肌收缩加强时,心缩期血流量所占的比例更小。由此可见,动脉舒张压的高低和心舒期的长短是影响冠脉血流量的重要因素。体循环外财阻力增大时,动脉舒张压升高,冠脉血流量增多。心率加快时,由于心动周期的缩短主要是心舒期缩短,故冠脉血流量也减少。右心室肌肉比较薄弱,收缩时对血流的影响不如左心室明显。在安静情况下,右心室收缩期的血流量和舒张期的血流量相差不多,或甚至多于后者。
(三)冠脉血流量的调节
对冠脉血流量进行调节的各种因素中,最重要的是心肌本身的代谢水平。交感和副交感神经也支配冠脉血管平滑肌,但它们的调节作用是次要的。
图4-29 一个心动周期中左、右冠状动脉血流变化情况(1mmHg=0.133kPa)
1.心肌代谢水平对冠脉血流量的影响 心肌收缩的能量来源几乎唯一地依靠有氧代谢。心肌因连续不断地进行舒缩,故耗氧量较大,即使在人体处于安静状态时,动脉血流经心脏后,其中65%-75%的氧被心肌摄取。因此心脏的动脉血和静脉血的含氧量差很大,换句话说,心肌提高从单位血液中摄取氧的潜力较不小。在肌肉运动、精神紧张等情况下,心肌代谢活动增强,耗氧量也随之增加。此时,机体主要通过冠脉血管舒张,即增加冠脉血流量来满足心肌对氧的需求。实验证明,冠脉血流量是和心肌代谢水平成正比的。在没有神经支配和循环激素作用的情况下,这种关系仍旧存在。目前认为,心肌代谢增强引起冠脉血管舒张的原因并非低氧本身,而是由于某些心肌代谢产物的增加。在各种代谢产物中,腺苷可能起最重要的作用。当心肌代谢增强而使局部组织中氧分压降低时,心肌细胞中的ATP分解为ADP和AMP。在冠脉血管周围的间质细胞中有5′-核苷酸酶,后者可使AMP分解产生腺苷。腺苷具有强烈的舒张小动脉的作用。腺苷生成的后,在几秒钟内即被破坏,因此不会引起其它器官的血管舒张。心肌的其它代谢产物如H+、CO2乳酸等,虽也能使冠脉舒张,但作用较弱。此外,缓激肽和前列腺素E等体液因素也能使冠脉血管舒张。
2.神经调节 冠状动脉受迷走神经和交感神经支配。迷走神经兴奋对冠状动脉的直接作用是引起舒张。但迷走神经兴奋又使心率减慢,心肌代谢率降低,这些因素可抵消迷走神经对冠状动脉的直接舒张作用。在动物实验中,如果使心率保持不变,则刺激迷走神经引起冠脉舒张。刺激心交感神经时,可激活冠脉平滑肌的α肾上腺素能受体,使血管收缩,但交感神经兴奋又同时激心肌的β肾上腺素能受体,使心率加快,心肌收缩加强,耗氧量增加,从而使冠脉舒张。给予β肾上腺素能受体拮抗剂后,刺激交感神经表现出直接的冠脉收缩反应。冠脉平滑肌上也有β肾上腺素能受体,后者被激活时引起冠脉舒张。交感神经兴奋对冠的β肾上腺至少能受体的激动一般不很明显。一些药物如异丙基肾上腺素对冠脉β肾上腺素能受体作用明显。
总之,在整体条件下,冠脉血流理主要是由心肌本身的代谢水平来调节的。神经因素对冠脉血流的影响在很短时间内就被心肌代谢改变所引起的血流变化所掩盖。
3.激素调节肾上腺素和去甲肾上腺素可通过增强心肌的代谢活动和耗氧量使冠脉血流量增加;也可直接作用于冠脉血管α或β肾上腺素能受体,引起冠脉血管收缩或舒张。甲状腺素增多时,心肌代谢加强,耗氧量增加,使冠状动脉舒张,血流量增加。大剂量血管升压素使冠状动脉收缩,冠脉血流量减少。血管紧张素Ⅱ也能使冠状动脉收缩,冠脉血流量减少。
二、肺 循 环
肺循环的功能是使血液在流经肺泡进和肺泡之间进行气体交换。呼吸性小支气管以上的呼吸道组织的营养物质收体循环的支气管的末梢之间有吻合支沟通。因此,有一部分支气管静脉血液可经过这些吻合支进入静脉和左心房,使主动脉血液中掺入1%-2%的静脉血。
(一)肺循环的生理特点
右心室的每分输出量和左心室的基本相同。肺动脉及其分支都较粗,管壁较主动脉及其分支薄。肺循环的全部血管都在胸腔内,而胸腔内的压力低于大气压。这些因素使肺循环有与体循环不同的一些特点。
1.血流阻力和血压 肺动脉管壁厚度仅为主动脉的三分之一,其分支短而管径较粗,故肺动脉的可扩张性较高,对血流的阻力较小。肺循环动脉部分总的阻力和静脉部分总的阻力大致相等,故血流在动脉部分的压力降落和在静脉部分的压力降落相等。肺循环毛细血管压大致在右心室压和左心房压数值的中点。由于肺循环血管对血流的阻力小,所以,虽然右心室的每分输出理和左心室每分输出量相等,但肺动脉压远较主动脉压为低。右心室压和肺动脉压可用插入导管的方法直接测量。在正常人,右心室收缩压平均约2.9kPa(22mmHg),舒张压为0-0.13kPa(0-1mmHg)。肺动脉的收缩压和右心室收缩压相同,平均为2.2kPa(22mmHg),舒张压为1.1kPa(8mmHg),平均压约1.7kPa(13mmHg)。用间接方法可测得肺循环行细血管平均压为0.9kPa(7mmHg)。肺循环的终点,即肺静脉和左心房内压为0.13-0.53kPa(1-4mmHg),平均约0.27kPa(2mmHg)。
2.肺的血容量 肺部的血容量约为450ml,占全身血量的9%。由于肺组织和肺血管的可扩张性大,故肺部血容量的变化范围较大。在用力呼气时,肺部血容量减少至约200ml;而在深吸气地可增加到约1000ml。由于肺的血容量较多,而且变化范围较大,故肺循环血管起着贮血库的作用。当机体失血时,肺循环可将一部分血液转移至体循环,起代偿作用。在每一个呼吸周期中,肺循环的血容量也发生周期性的变化,并对左心室输出量和动脉血压发生影响。在吸气时,由腔静脉回流入右心房的血量增多,右心室射出的血量也就增加。由于肺扩张时可将肺循环的血管牵拉扩张,使其容量增大,能容纳较多的血液而由肺静脉回流入左心房的血液则减少。但在几次心搏后,扩张的肺循环血管已被充盈,故肺静脉回流入左心房的血量逐渐增加。在呼气时,发生相反的过程。因此,在吸气开始时,动脉血压下降,到吸气相反相的后半期降至最低点,以后逐渐回升,在呼气相的后半期达到最高点。在呼吸周期中出现的这种血压波动,称为动脉血压的呼吸波。
3.肺循环毛细血管外的液体交换 如前所述,肺循环毛细血管平均约0.9kPa(7mmHg),而血浆胶体渗透压平均3.3kPa(25mmHg),故将组织中的液体吸收入毛细血管的力量较大。现在一般认肺部组织液的压力为负压。这一负压使肺泡膜和毛细血管管壁互相紧密相贴,有利于肺胞和血液之间的气体交换。组织液负压还有利于吸收肺泡内的液体,使肺泡内没有液体积聚。在某些病理情况下,如左心衰竭时,肺静脉压力升高,肺循环毛细血管压也随着升高,就可使液体积聚在肺泡或肺的组织间隙中,形成肺水肿。
(二)肺循环血流量的调节
1.神经调节 肺循环血管受交感神经和迷走神经支配。刺激交感神经对肺血管的直接作用是引起收缩和血流阻力增大。但在整体情况下,交感神经兴奋时体循环的血管收缩,将一部分血液挤入肺循环,使肺循环内血容量增加。循环血液中的儿茶酚胺也有同样的效应。刺激迷走神经可使肺血管舒张。乙酰胆碱也能使肺血管舒张,但在流经肺部后即分解失活。
2.肺泡气的氧分压 肺泡气的氧分压对肺部血管的舒缩活动有明显的影响。急性或慢性的低氧都能使肺部血管收缩,血流阻力增大。引起肺血管收缩的原因是肺泡气的氧分压低而不是血管内血液的氧张力低。当一部分肺泡内气体的氧分压低时,这些肺泡周围的微动脉收缩。在肺泡气的CO2分压升高时,低氧引起的肺部微动脉的收缩更加显著。可见肺循环血管对局部低氧发生的反应和体循环血管不同。肺部血管对低氧发生缩血管反应的机制,目前还不完全清楚。有人推测低氧可能使肺组织产生一种缩血管物质,也有人认为必需有血管内皮存在才能发生这种缩血管反应。肺泡氯低氧引起局部缩血管反应,具有一定的生理意义。当一部分肺泡因通气不足而氧分压降低时,这些肺泡周围的血管收缩,血流减少,而使较多的血液流经通气充足,肺泡气氧分压高的肺泡。假如没有这种缩血管反应,血液流经通气不足的肺泡时,血液不能充分氧合,这部分含氧较低的血液回流入左心房,就会影响体循环血液的含氧量。当吸入气氧分压过低时,例如在高海拔地区,可引起肺循环动脉广泛收缩,血流阻力增大,故肺动脉压显著升高。长期居住在高海拔地区的人,常可因肺动脉高压使右心室负荷长期加重而导致右心室肥厚。
3.血管活性物质对肺血管的影响 肾上腺素、去甲肾上腺素、血管紧张素Ⅱ、血栓素A2、前列腺素F2α等能使肺循环的微动脉收缩。组胺、5-羟色胺能使肺循环静脉收缩,但在流经肺循环后即分解失活。
三、脑 循 环
脑组织的代谢水平高,血流量较多。在安静情况下,每百克脑的血流量为50-60ml/min。整个脑的血流量约为750ml/min。可见,脑的比重虽仅占体重的约2%,但血流量却占心输出量的15%左右。脑组织的耗氧量也较大。在安静情况下,每百克脑每分钟耗氧3-3.5ml;或者说,整个脑的耗氧量约占全身耗氧量的20%。
(一)脑循环的特点
脑位于颅腔内。颅腔是骨性的,其容积是固定的。颇腔内为脑、脑血管和脑脊液所充满,三者的容积的总和也是固定的。由于脑组织是不可压缩的,故脑血管舒缩程度受到相当的限制,血流量的变化较其它器官的为小。
脑循环的毛细血管壁内皮细胞相互接触紧密,并有一定的重叠,管壁上没有小孔。另外,毛细血管和神经元之间并不直接接触,而为神经胶质细胞怕隔开。这一结构特征对于物质在血液和脑组织之间的扩散起着屏障的作用,称为血-脑屏障(blood-brain barrier)。
(二)脑血流量的调节
1.脑血管的自身调节 脑血流量取决于脑的动、静脉的压力差和脑血管的血流阻力。在正常情况下,颈内静脉压接近于右心房压,且变化不大,故影响血流量的主要因素是颈动脉压。政党情况下脑循环的灌注压为10.6-13.3kPa(80-100mmHg)。平均动脉压降低或颅内压升高都可以使脑的灌注压降低。但当平均动脉压在8.0-18.6kPa(60-140mmHg)范围内变化时,脑血管可通过自身调节的机制使脑血流量保持恒定。平均动脉压降低到8.0kPa(60mmHg)以下时,脑血流量就会显著减少,引起脑的功能障碍。反之,当平均动脉压超过脑血管自身调节的上限时,脑血流量显著增加。
2.CO2和O2分压对及脑血流量的影响血液CO2分压升高时,脑血管舒张,血流量增加。CO2过多时,通过使细胞外液H+浓度升高而使脑血管舒张。过度通气时,CO2呼出过多,动脉血CO2分压过低,脑血流量减少,可引起头晕等症状。血液O2分压降低时,也能使脑血管舒张。
3.脑的代谢对脑血流的影响 脑的各部分的血流量与该部分脑组织的代谢活动程度有关。实验证明,在同一时间内脑的没部分的血流量是不同的,当脑的某一部分活动加强时,该部分的血流量就增多。例如在握拳时,对侧大脑皮层运动区的血流量就增加;阅读时脑的许多区域血流量增加,特别是皮层枕叶和颞叶与语言功能有关的部分血流量增加更为明显。代谢活动加强引起的局部脑血流量增加的机制,可能是通过代谢产物如H+离子、K+离子、腺苷,以及氧分子降低,引起脑血管舒张的。
4.神经调节 颈上神经节发出的去甲云彩上腺素后纤维,其末梢分布至脑的动脉和静脉,并分布至软脑膜的血管,还有少量分布至脑实质的血管。脑实质内的小血管有起自蓝斑去甲肾上腺素神经元的轴突末梢的分布。副交感乙酰胆碱能神经末梢也分布至脑血管。此外,脑血管政治家血管活性肠肽等神经肽纤维末梢分布。神经对脑血管活动的调节作用不很明显。刺激或切除支配脑血管的交感或副交感神经,脑血流量没有明显变化。在多种心血管反射中,脑血流量一般变化都很小。
(三)脑脊液的生成和吸收
脑脊液存在于脑室系统、脑周围的脑池和蛛网膜下腔内,可被视为脑和脊髓的组织液和淋巴。成年人的脑脊液总量约150ml。每天生成的脑脊液约800ml,为脑脊液总量的5-6倍。但同时有等量的脑脊液被吸收入血液,可见脑脊液的更新率较高。
脑脊液主要由侧脑室、第三脑室和第四脑室的脉络丛分泌。侧脑室内的脑脊液经室间孔流入第三脑室,再经过导水管进入第四脑室,然后进入蛛网膜下腔。除脉丛外,室管膜细胞也能分泌脑脊液。软脑膜血管和脑的毛细血管滤过的液体,一部分被重吸收,其余的则沿着血管周围间隙进入蛛网膜下腔,成为脑脊液的一部分。
脑脊液主要通过蛛网膜绒毛被吸收入静脉的血液内。蛛网膜绒毛有活瓣状的细微的管道,其直径为4-12μm。当蛛网膜下腔的压力高于静脉窦的压力时,这些管道就开放。这时,脑脊液(包括其中所含的蛋白质分子甚至小的颗粒如红细胞等)可进入静脉窦血液。当蛛网膜下深的压力低于静脉窦压力时,管道关闭,液体不能由静脉窦向蛛网膜下腔倒流。脑脊液压力的高低取决于其生成和吸收之间的平衡关系。正常人在到卧位时,脑脊液压平均为1.3kPa(10mmHg)。当脑脊液有吸收受到阻碍时,脑脊液压就会升高,并影响脑血流和脑的功能。
脑脊液的主要功能是在脑、脊髓和颇腔、椎管之间起缓冲的作用,有保护性意义。脑浸浴于脑脊液中,由于浮力的作用,使脑的重量减轻到仅50g左右。另外,脑脊液还作为脑和血液之间进行物质交换的中介。脑组织中没有淋巴管,由毛细血管漏出的少量蛋白质,主要经过血管周围间隙进入蛛肉膜下腔的脑脊液中,然后通过蛛网膜绒毛回入血液。
(四)血-脑脊液屏障和血-脑屏障
脑脊液主要是由脉络丛分泌的,但其成分和血浆不脑脊液中蛋白质的含量极微,葡萄糖含量也较血浆为少,但Na+ 和Mg2+的浓度较血浆中的高,K+、HCO3-和Ca2+的浓度则较血浆中的低。可见,血液和脑脊液之间物质的转运并不是被动的过程,而是主动转运过程。另外,一些大分子物质较难从血液进入脑脊液,仿佛在血液和脑脊液之间存在着某种特殊的屏障,故称之为血-脑脊液屏障(blood-cerebrospinal fluid barrier)。这种屏障对不同物质的通透性上不同的。例如O2、CO2等脂溶性物质可很容易地通过屏障,但许多离子的通透性则较低。血-脑脊液屏障的基础是无孔的毛细血管壁和脉络丛细胞中运输各种物质的特殊载体系统。
血液和脑组织之间也存在着类似的屏障,可限制物质在血液和脑组织之间的自由交换,称为血-脑屏障。脂溶性物质如O2、CO2、某些麻醉药以及乙醇等,很容易通过血-脑屏障。对于不同的水溶性物质来说,其通透性并不一定和分子的大小相关。例如葡萄糖和氨基酸的通透性较高,而甘露醇、蔗糖和许多离P嬀通透性则很低,甚至不能通透。这说明脑内毛细血管处的物质交换和身体其它部分的毛细血管处是不同的,也是一种主动的转运过程。用电子显微镜观察,脑内大多数毛细血管表面都被星状胶质细胞伸出的突起(血管周足)所包围。因此推测,毛细血管的血液和神经元之间的物质交换可能都要通过胶质细胞作为中介。因此,毛细血管的内皮、基膜和星状胶质细胞的血管周足等结构可能是血-脑屏障的形态学基础。另外,毛细血管壁对各种物质特殊的通透性也和这种屏障作用有重要的关系。
血-脑脊液屏障和血-脑屏障的存在,对于保护脑组织周围稳定的化学环境和防止血液中有害物质侵入脑内具有重要的生理意义。例如,脑脊液中K+的浓度较低,即使在实验中使血浆K+浓度加倍,脑脊液中K+浓度仍能保持在正常水平。因此脑内神经元的兴奋性不会因血浆中K+浓度的变化而发生明显的变化。由于血-脑屏障的存在,循环血液中的乙酰胆碱、去甲肾上腺素、多巴胺、甘氨酸等物质就不易进入脑,否则,血浆中这些物质浓度的改变将会明显地扰乱脑内神经元的政党功能活动。
需要指出,脑的某些部分,如下丘脑第三脑室周围和延髓后缘区等处的室周器官,血-脑屏障比较薄弱,毛细血管壁对许多物质的通透性高于脑的其它部分。因此循环血液中的有些物质,如血管紧张素Ⅱ和其它肽类物质,可以在这些部位进入脑内,作用于相应的受体,引起各种效应。另外,当脑组织发生缺氧、损伤等情况以及在脑肿瘤部位,毛细血管壁的通透性增加,故平时不易透过血-脑屏障的物质进入受损部位的脑组织。在临床上可以用同位素标记的白蛋白注入体内,这些蛋白质进入正常脑组织的速度很慢,但较易进入脑肿瘤组织,因此可用这种方法匀来检查脑瘤的部位。在用药物治疗神经系统疾病时,必须明确所用的药物是否容易通过血-脑屏障。
在脑室系统,脑脊液和脑组织之间为室管所分隔;在脑的表面,脑脊液和脑组织之间为软脑膜所分隔。室管膜和软脑膜的通透性很高,脑脊液中的物质很容易通过室管膜或软脑膜进入脑组织。因此,在临床上可将不易通过血-脑屏障的药物直接注入脑脊液,使之能较快地进入脑组织。
参考资料
1.徐丰彦,张镜如主编,人体生理学。北京:人民卫生出版社,1989
2.何瑞荣主编,心血管生理学 北京:人民卫生出版社,1987
3.骆鸿,何瑞荣,延髓腹外侧部对心血管活动的调节。生理科学进展1987;18:224-229
4.姚泰,中枢神经系统对血压的调节,生理科学进展,1989;20:276-283
5.韩济生主编,神经科学纲要,第47章,北京:北京医科大学、中国协和医科大学联合出版社,1983
6.Little RC.Physiology of the Heart and Circulation 3rd ed,Year Book Medical Publishers,In,Chica-go,1985
7.Fozzard HA et al.The Heart and Cardiovascular Systen-Scientific Foundations,Raven Press,New York,1986
8.Noble D.The surprising teart, a revieq of recent progress in cardiac electrophysiology. J Physiol 1984;353:1-50
9.Reuter H.Ion channels in cardiac cell menbrane.Ann Rev Physiol 1984;46:473-484
10.Irisawa H et al. Pacemaker currents in mammalian nodal cells .J Mol cell Cardiol 1984;16:777-781
11.Winegrad S.Regulation of cardiac contractle proteins.Circ Res 1984;55:565-574
12.Siegl PKS.Overview of cardiac inotropic mechanisms.J Cardiovasc Pharmacol 1986;8(suppl 9):S1-S10
13 .Carafoli E Intracellular calcium homeostasis.Ann Rev Biochem 1987;56:395-433
14.Brown JH et al.Phosphoinositide-generated second messengers in cardiac signal transduction.Trends in Cardiovascular Medicine 1922;2:199-213
15.Ballermann VJ,Zeidel ML,Gunning ME,Brenner BM,Vasoactive piptides and the kidney.in Kidney 4th ed, Brenner BM, Rector Jr FC ed,Chapter 14,Wb Saunders Co,Philadelphia,1991
16.Cowley Jr AW,Long-term control of arterial blood pressure.Physiol Rev 1992;72:231-300
17.Guyton AC.Textbook of Medical Physiology 8th ed,Wb Saunders Co,Philadelphia,1991
18.Johnson LR.Essential Medical Physiology,Raven Press,New York,1992
19.Mione MC,Ralevic V,Burnstock G.Peptdes and vasomotor mechanisms.Pharmac Ther 1990;46:429-468
20 Patton HD,Fuchs AF,Hille B,Scher AM,Steiner R.Textbook of Physiology 21sted,WB Saunders Co,Philadelphia,1989