
���Ľڡ���ϸ������������
�������������ʽ���˶�����Ҫ�ǿ�һЩ��ϸ�������������ɵġ����磬����ĸ����˶��ͺ��������ɹ���������������ɣ��������Ѫ����ļ�����������ɣ�һЩ�п�������θ�������ס��ӹ���Ѫ�ܵ����ٵ��˶�������ƽ��������������ɡ���ͬ������֯�ڹ��ܺͽṹ�ϸ����ص㣬���ӷ���ˮƽ�������������������ϸ�������������������ʣ���Ҫ�뼡�����ͼ��˵���������йأ����������Ź��̵Ŀ��ƣ�Ҳ��ijЩ����֮�����������о����ֵĹ�����Ϊ�ص㣬˵����ϸ�����������ơ�
����������������������֯��Լռ���ص�40%���ڹǺؽڵ�����£�ͨ�������������������ţ�����˺ߵȶ���ĸ��������˶����������ɴ��������ļ���ά��ɣ�ÿ������ά����һ����ϸ�������˼���ά��ϸ��Բ���Σ�ֱ��Լ60��m�����ɴ�������������ʮ���ס��ڴ���������У������ͼ���ά����ƽ�����У��������˶����ɽ����֯���ɵ������ںϣ����߸����ڹ��ϣ�ͨ����֫�Ĺ������ڸ��ŵ�֮������Ҫ���һ���ؽڣ�ͨ����������������ţ��Ϳ�������֫�����������ֱ�����ǵ������Ͷ�������������ȣ�����������������ϵĻ�Ľ����ÿ����������ά����һ�������Ĺ��ܺͽṹ��λ���������ٽ���һ���˶���ĩ�ҵ�֧�䣬���������������άֻ����֧�����ǵ�����ά���嶯����ʱ�����ܽ�����������ˣ��������еĹ������������������ϵͳ�Ŀ�������ɵġ�
����һ����-��������ͷ�����˷ܴ���
�����˶�����ά�ڵ�����ĩ�Ҵ�ʱ��ʧȥ���ʣ�����¶����ͻĩ��Ƕ�뵽��ϸ��Ĥ�ϳ����հ��Ĥ�����У�����ͻĩ�ҵ�Ĥ���հ�Ĥ����ֱ�ӽӴ������DZ�������ϸ����Һ�Ľ�ͷ��϶�����������к��гɷֲ����Ļ��ʣ���ʱ��ĩ���·����հ�Ĥ���й��������ϸ���ڰ��룬�γ��������ޣ�����������������ӽ�ͷ��Ĥ�������ʹ���������ɽ϶���Ŀ�ĵ����ʷ��ӣ������������ΪN-�������������壬����֤��������һЩ��ѧ�ſ�ͨ������������ACh�����Խ�ϵ��ǵ�λ������ͻĩ�ҵ��Ὤ�У���������������������д���ֱ��Լ50nm�������������ݣ�ͼ2-19��������֯��ѧ�ķ�������֤���������ں���ACh����ACh�������Ὤ�кϳɣ�Ȼ�������������ڡ��ݲⶨ��ÿ�������������ACh��ͨ�����൱�㶨�ģ��ҵ����DZ��ͷ�ʱ��Ҳ��ͨ���������ã�������Ϊ��λ�����ҡ��ͷţ�����Ϊ����ʽ�ͷš�����ĩ�Ҵ��ڰ���״̬ʱ��һ��ֻ��������������ؽ����ͷţ����ܶԼ�ϸ����������Ӱ�졣������ĩ�Ҵ����嶯����ʱ���ڶ�����λ��ɵľֲ�Ĥȥ������Ӱ���£�������������ͻĤ���ڲ��濿����ͨ������Ĥ����ͻĤ���ںϣ������ںϴ������ѿڣ�ʹ�����е�AChȫ�������ͷ��϶�������㣬һ�ζ�����λ�ĵ����ʹ��Լ200��300�����ݵ������ŷţ�ʹ��107��ACh���ӱ��ͷš���ͻĩ�Ҵ��ĵ�λ�仯���������ŷŵĹ���ʮ�ָ��ӣ�����������ͻĩ��Ĥ��ȥ�����������˸ô����еĵ�ѹ�ſ�ʽCa2+ͨ�����ţ�����ϸ����϶Һ�е�Ca2+������ͻĩ�ң������������ƶ������ŷŵĹ��̡�Ca2+�Ľ������ƺ������������ͷŵ���Ŀ��ϸ����Һ�е�Ca2+�ͣ���Mg2+�������谭ACh���ͷŶ�Ӱ����-����ͷ���������ܡ��ѹʷ����Ժʿ��30�������-����ͷ�Ļ�ѧ���ʴ��ݽ��й���Ҫ���о���
ͼ2-19 ��-����ͷ���ij��ṹʾ��ͼ
������ACh����ͨ����ͷ��϶�����հ�Ĥ����ʱ������ͬ���д����ڸô�������ͨ�������ʵ�������-�ǵ�λ��ϣ�ÿ���ӵ�ͨ��������������ӵ�ACh���ɴ�����ĵ����ʷ����ڲ�����ı仯�ᵼ������ͨ���ṹ�Ŀ��š�����ͨ������ʱ�����ĺ�����ǰ���ᵽ��Na+ͨ�������Ϊ������Na+��K+��������Ca2+ͬʱͨ���������⼸����������ʱ��Ĥ�ڴ��ķֲ��ص㣬ʵ�ʳ��ֵ���Na+��������K+�����������ܵĽ����ʹ�հ�Ĥ��ԭ�о�Ϣ��λ��С������ֵ�������༴����Ĥ��ȥ��������һ��仯����Ϊ�հ��λ�����ij���Լ���嶯�����ͷǰĤ����0.5��1.0ms�����������˶����嶯����ĩ��ʱ����¼�����ڸ������ݵ��Է��ͷ����հ�Ĥ�������С�ĵ�仯����Ϊ�հ��λ���հ��λ��ǰ���ľֲ��˷ܵ練Ӧ�����Ƶ����ʣ������֡�ȫ���ޡ����ԣ����С���ͷǰĤ�ͷŵ�ACh�����ɱ�������Ӧ�ڣ��ɱ����ܺ�����ȣ�������һ���¼�����հ��λ���Ƕ����հ��λ�ܺ͵Ľ�����հ��λ����ʱ�������Ե��������������ʽӰ���հ�Ĥ��Χһ�㼡ϸ��Ĥ��һ��ļ�ϸ��Ĥ������ͻ��Ĥ�������ƣ�������Ҫ����ѹ�ſ�ʽNa+ͨ����K+ͨ���������ͬ�հ�Ĥ�ڽӵļ�ϸ��Ĥ�ľ�Ϣ��λ�����հ��λ��Ӱ���ȥ�������ô�Ĥ���е�λˮƽʱ���ͻ�����һ����������ϸ��Ĥ����ȫ���ޡ�ʽ�����Ķ�����λ��������ͨ����ν���˷�-����������������ϸ������һ�λ�е������
������������£�һ���嶯���ͷŵ�ACh�Լ�����������հ��λ�Ĵ�С����Լ��������ϸ��Ĥ������λ������ֵ��3��4�����������ͷ�����˷ܴ���ͨ����1��1�ģ��༴�˶���άÿ��һ���嶯����ĩ�ң����ܡ��ɿ��ء�ʹ��ϸ���˷�һ�Σ��շ�һ����������һ���뽫��Ҫ������Ԫ֮����˷ܴ��������Բ�ͬ������ʮ�£�����ͷ�����ܱ���1��1�Ĺ�ϵ����Ҫ��ÿһ���嶯���ͷŵ�ACh�ܹ���������һ�μ����˷ܺ�Ѹ��������������������������հ��ʹ�հ�Ĥ����ȥ��������Ӱ���´ε������嶯��ЧӦ����֪��ACh�������Ҫ��������ø�Ľ�����������ɣ���ø��Ҫ�ֲ��ڽ�ͷ��϶�кͽ�ͷ��Ĥ�ϣ����Ǵ�Լ������2.0ms��ʱ���ڽ�һ���嶯���ͷŵ�ACh�����������ҩ����������ڽ�ͷ���ݹ����еIJ�ͬ�Σ�Ӱ�������Ľ�ͷ���ܡ����磬�������ͦ�-�����߶�����ͬACh�����հ�Ĥ��ACh�����ǵ�λ�����������Ͻ�ͷ���ݶ�ʹ����ʧȥ�������������������õ�ҩ���Ϊ�����ɳڼ����л���ũҩ����˹�����Ե�����ø��ѡ���Ե��������ã������ACh�ڽ�ͷ��������λ�Ĵ������ۣ����������ж�֢״����֢���������������ڹ������հ崦��ACh�ſ�ͨ��������������ϰ�������
��������������ϸ����ϸ�ṹ
����������ϸ���ڽṹ����ͻ��֮�㣬�Ǻ��д����ļ�ԭ��ά�ͷḻ�ļ���ϵͳ���������и߶ȹ�������ϸ�������ں�����������ɻ�����ֻ�е�˶��Ĺ��ܵ�λ��
������һ����ԭ��ά�ͼ�С��
����ÿ������ά���д���ֱ��1��2��m����ά״�ṹ����Ϊ��ԭ��ά������ƽ�����У��ݹἡ��άȫ������һ��ϸ���пɴ���ǧ��֮�ࣨͼ2-20����ÿ����ԭ��ά��ȫ�������ֹ�������������棬�ֱ��Ϊ�����Ͱ�����������ƽ�еĸ���ԭ��ά֮�䣬�����Ͱ����ֶ��ֲ���ͬһˮƽ�ϣ������ij��ȱȽϹ̶������ۼ���ھ�ֹ���ܵ�����ǣ�����������ʱ����������1.5��m�ij��ȣ��ڰ������룬��һ�������������ΪH�������ij����漡������״̬�IJ�ͬ���б仯����H�������༴�������������룬����һ������İ��ߣ���ΪM�ߡ������ij����ǿɱ�ģ����ڼ��ⰲ��ʱ�ϳ���������һ����Χ�ڿ��������ܵı�ǣ�����䳤���������ڼ�������ʱ�ɱ�̡���������Ҳ��һ������İ��ߣ���Ϊz�ߣ���z�̣���Ŀǰ�Ѿ��϶�����ԭ��ά��ÿһ��λ������z��֮��������Ǽ������������ŵ��������λ��������һ��λ���м䲿�ֵİ����������1/2���������ϳ�Ϊ��С�ڣ�sarcomere�������������ij��ȿɱ䣬��С�ڵij����ڲ�ͬ����¿ɱ䶯��1.5��3.5��m֮�䣻ͨ���������������ʱ��С�ڵij���ԼΪ2.0��2.2��m��

ͼ2-20 ������ϸ���ļ�ԭ��ά�ͼ���ϵͳ
ע�⼡С���ܵ���ɺͲ�ͬ��λ��С�ں�ϲ��֡�ϸ��˿�ļ�������
������X-������ȸ����ܵķ�����һ�����֣���С�ڵ������Ͱ��������и�ϸ�ġ�ƽ�����е�˿״�ṹ����Ϊ��˿�������к��еļ�˿�ϴ֣�ֱ��Լ10nm����Ϊ�ּ�˿���䳤���밵����ͬ��ʵ���ϰ������γɾ������ڴּ�˿�Ĵ��ڣ�M�����ǰѳ����Ĵּ�˿�̶���һ��λ�õ�ij�ֽṹ�������еļ�˿��ϸ��ֱ��Լ5nm����Ϊϸ��˿��������z�߽ṹ���������������ÿ��ij��ȶ���1.0nm������������ڼ�С���ܳ���С��3.5nm������£���Ȼ��һ��Ҫ���밵�����ʹּ�˿���ڽ������ص���״̬�����������z�����밵����ϸ��˿δ������������һ�ξ��룬����γ���H�������ⱻ����ʱ����С�ڳ���������ʱϸ��˿�ɰ����ص���������ʹ������������H��Ҳ��Ӧ������ͼ2-20���£���
�����֡�ϸ��˿��ص�ʱ���ڿռ���Ҳ���ֹ������У���ɴӼ�ԭ��ά�ĺ�����Ͽ�����ͼ2-20�·���ʾ�ڼ�С�ڵIJ�ͬλ�ý���ԭ��ά���ʱ�����������ܿ��������ּ�˿�ķֲ��������ͨ�������ĺ������ֻ��ϸ��˿�����ǵ�����λ���൱��һ���������εĸ����㣻��ͨ��H���ĺ������ֻ�дּ�˿�����Ƕ������������εĸ������ϣ�����H������İ����ĺ�����ϣ���ɿ����֡�ϸ��˿�������ڵ���������ÿһ���ּ�˿������������ϸ��˿Ϊ������������ε����룬�����Ϊ����ʱ��ϸ��˿֮������������������
��������������ϵͳ
��������ϵͳָ������ÿһ����ԭ��ά��Χ��Ĥ���ҹ�״�ṹ������Դ���ܶ�����ͬ����������Ĺܵ�ϵͳ��ɡ�һ���ּ��ܵ����з���ͼ�ԭ��ά�ഹֱ����Ϊ���ϵͳ���T�ܣ����ɼ�ϸ���ı���Ĥ���ڰ�����γɣ����Ǵ����ڼ�ԭ��ά֮�䣬����z��ˮƽ����Щ�������ڰ����������νӴ���ˮƽ���γɻ��Ƽ�ԭ��ά�Ĺܵ����������ͨ����ǻͨ����Ĥ���봦��С����ϸ����Һ��ͨ�����������뵽ϸ���Ľ�ԡҺ�У���Щ���ʿ��Ժܿ���ÿһ�����Ƽ�С�ڵĺ��ϵͳ�г��֣������ܽ��뼡���ͼ�������ȥ����ԭ��ά��Χ������һ�鼡��ϵͳ�����Ǽ����������ǵ����з���ͼ�С��ƽ�У���Ϊ�ݹ�ϵͳ���ΪL�ܣ��ݹ�ϵͳ������Ҫ����ÿ����С�ڵ��м䲿�֣�����һЩ���ͨ�Ĺܵ��������ڽӽ���С�����˵ĺ��ʱ��ǻ�������Ϊ��ĩ�أ���ʹ�ݹ��Խϴ������ͺ�������ÿһ��ܺ��������༡С�ڵ��ݹ���ĩ�أ������������ܽṹ��ͼ2-20���ϣ������о�����ܺ��ݹܵ�Ĥ�������ܽṹ�������Ӵ����м��и���Լ12nm�ļ�϶��˵������ܵ�����ǻ����ֱ�ӹ�ͨ���������Ľṹ��Ȼ������ϸ������֮��ij����ʽ����Ϣ���ݡ�Ŀǰ�ձ���ϵĿ����ǣ����ϵͳ�������ǽ���ϸ���˷�ʱ������ϸ��Ĥ�ϵĵ�仯��T��Ĥ����ϸ���ڲ�������������ĩ�ص�������ͨ���Ը����ӵ����桢�ͷź��ٻ��ۣ�������С�ڵ����������ţ��������ܽṹ�ǰѼ�ϸ��Ĥ�ĵ�仯��ϸ���ڵ����������νӻ����������Ĺؼ���λ��
�����������������������ƺ��˷�-��������
����Huxley����50������ھ�������ü�С���д֡�ϸ��˿���������˵�����������Ļ��ơ���һ����Ϊ�������ۣ�sliding theory������Ҫ�����ǣ���������ʱ��Ȼ������Ͽ��Կ��������������ά�����̣����ڼ�ϸ���ڲ���˿�����������ķ��ӽṹ�����̣���ֻ����ÿһ����С���ڷ�����ϸ��˿��ּ���֮��Ļ��У��༴��z�߷�����ϸ��˿��ij�������������������������ƶ�����������ڵ�z�߶����������С�ڳ��ȱ�̣����������ԭ��ά����ϸ�������������ⳤ�ȵ����̡�����������ֱ�ӵ�֤���ǣ���������ʱ���ް������ȵı仯����ֻ�ܿ����������ȵ����̣��������ͬʱҲ������������H����Ӧ�ر�խ����ֻ��˵����ϸ��˿�ڼ�������ʱҲû�����̣�ֻ�����Ǹ��������ƶ����ʹּ�˿�����˸���̶ȵ��ص������ֱ仯ֻ���ô֡�ϸ��˿֮�����������˶����������������͡�����������Ҫ��һ��˵���������ǣ���������ʱ������ʲô������ʹϸ��˿��ּ�˿֮�们�У��Լ���������Щ���̺ͼ���Ĥ���˷ܹ�����ϵ�����������������ڼ������ﻯѧ������ϸ������ѧ�����ķ�չ����˿���еĻ����ѻ����ϴ���ɼ�˿�ĵ����ʷ��ӽṹ��ˮƽ�õ������������뻬�еĿ�ʼ����ֹ�йصĸ��ֿ������أ�Ҳ���˽Ͻ�������˽⡣
������һ����˿�ķ�����ɺͺ��ŵ��˶�
���������������������ɼ�˿�ĵ����ʷ��ӽṹ�����ǵ�������ֱ�ӵĹ�ϵ���ּ�˿��Ҫ�ɼ������ף���Ƽ��ף�����ɣ����ǵķ����ڴּ�˿�гʶ��ص��й�������С�һ���ּ�˿��Լ����200��300�����������ӣ�ÿ�����ӳ�150nm���ʳ���״����һ������״�������ɴּ�˿ʱ������״������M�߶��ۺϳ������γɴּ�˿�����ɣ���״�����й������¶��M������Ĵּ�˿���ɵı��棬�γɺ��ţ�ͼ2-21���������ⰲ��ʱ�����������ɵķ����ഹֱ���ɴּ�˿����ͻ��Լ6nm����X-�����䷨֤���������ڴּ�˿����ķֲ�λ��Ҳ���ϸ��й���ģ����ڴּ�˿��ͬһ�ܾ���ֻ�����������180��ĺ���ͻ����������ܾ����14.3nm������������һ�Ժ���ͻ��������ǰһ����60��ļнǣ���˷����������ĶԺ��ų���ʱ���䷽���������һ�Ժ�����ƽ�У������һ�Ժ������42.9nm��ͼʾ-21���ң����������ŵķֲ������������һ���ּ�˿Ϊ6��ϸ��˿�����Ƶ�������Ӧ���༴�����к��ų��ֵ�λ�ã�������һ��ϸ��˿��֮��ԣ�������ÿ��ϸ��˿��˵���ּ�˿����ÿ��42.9nm�����һ��������֮��ԣ�ͼ2-21�������ֶ�Ӧ��ϵ�����ڴ֡�ϸ��˿֮����������Ȼ��ʮ�������ġ�
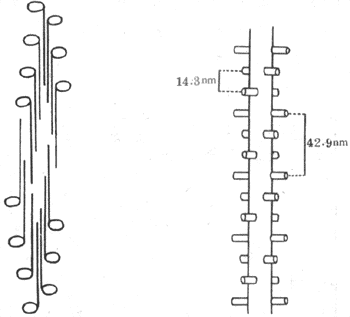
ͼ2-21 �ּ�˿�м��������ӵ�����ʾ��ͼ
���������ӵij���״������ۺϣ��γɴּ�˿���ɣ���״����¶�ڱ��棬�γɺ���
�ҡ������ڴּ�˿����ļ�������
��������֤�������������е����ﻯѧ���Զ��ڼ�˿�Ļ�������Ҫ���塣���ŵ���Ҫ�����ж���һ�Ǻ�����һ�������¿��Ժ�ϸ��˿�ϵļ��˵����ӳʿ����ԵĽ�ϣ�ͬʱ���ֺ�����M�߷����Ť�����̶����ֺ��ź�ϸ��˿�Ľ��롢��λ��Ȼ����ͬϸ��˿������ĵ��ϣ������µ�Ť������˷�����ʹϸ��˿������M�߷����ƶ������Ǻ��ž���ATPø�����ã����Էֽ�ATP�������������Ϊ���Űڶ���������������Դ���ɴ˿ɼ������ź�ϸ��˿������ã�������˿���еı�Ҫ������
����ϸ��˿���������ֵ�������ɣ�����60%�Ǽ��˵��ף���Ƽ������ף������˵����뼡˿������ֱ�ӵĹ�ϵ���ʺͼ�������һͬ����Ϊ���������ʡ����˵����ӵ������״����������ϸ��˿�оۺϳ�˫����״����Ϊϸ��˿�����ɣ�ͼ2-22����ϸ��˿�����������ֵ����ʣ����Dz�ֱ�Ӳ��뼡˿�������ã�����Ӱ��Ϳ�������������֮�������ã��ʳ�Ϊ���ڵ����ʣ�����һ����ԭ�������ף�Ҳ��˫�����ṹ����ϸ��˿�кͼ��˵���˫�������У����ڼ��ⰲ��ʱԭ��������λ�������ڼ��˵��ͺ���֮�䣨ͼ2-23������������谭�������ϵ����ã���һ�ֵ��ڵ����ʳ�Ϊ���Ƶ��ף����ԭ�����ף������Ƶ�����ϸ��˿�ϲ�ֱ�Ӻͼ��˵����������ӣ���ֻ����һ���ļ��������ԭ��������˫�����ṹ֮�ϡ����Ƶ��ķ��ӳ����Σ����������ǵ�λ��ͼ2-22�����ǵ�λC����һЩ��˫����ɵĽ��λ�㣬����Լ����г��ֵ�Ca2+���Լ��������ܳ��ֵ����������Ӻ�H+���кܴ���������ǵ�λT�����ǰ��������Ƶ����ӽ����ԭ�������ף����ǵ�λI�����������ǵ�λC��Ca2+���ʱ������Ϣ���ݸ�ԭ�������ף�������ߵķ��ӹ������ı䣬������Լ��˵��ͺ������ϵ��谭���á�
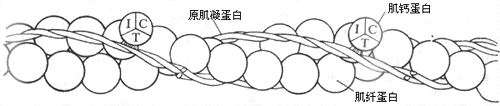
ͼ 2-22 ϸ��˿�ķ������
I��T��C�ֱ�������Ƶ��������ǵ�λ
������˿���еĻ�������һ�㹫��Ϊ������ϸ���ϵĶ�����λ������Ca2+Ũ������ʱ����ΪCa2+����ļ��Ƶ�������㹻������Ca2+����������˼��Ƶ����ӹ����ijЩ�ı䣬���ָı䡰���ݡ�����ԭ�������ף�ʹ���ߵĹ���Ҳ����ijЩ�ı䣬������ʹԭ��������˫�����ṹ������ij��Ťת����ͰѰ���ʱ��ֹ���˵��ͺ������ϵ��谭���س�ȥ�����������ߵĽ�ϡ��ں����뼡�˵��Ľ�ϡ�Ť����������ٽ�ϡ���Ť�����ɵĺ���ѭ�������У�ʹϸ��˿�����������ƶ��������������ATP�ķֽ����ĺͻ�ѧ�����е�ܵ�ת��������˼����������ͼʾ2-23����
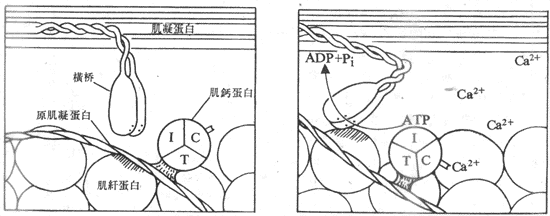
ͼ2-23 Ca2+ͨ���ͼ��Ƶ��Ľ�ϣ��շ����źͼ��˵���֮��������
���������ĺ���ѭ����һ����С���������������ж��Ƿ�ͬ���ؽ��еģ������ſ���ʹ��������㶨�����������������̡������ܲ���ѭ���ĺ�����Ŀ�Լ�����ѭ���Ľ������ʣ����Ǿ����������̶̳ȡ������ٶ��Լ������������Ĺؼ����أ��⽫�ڷ���������������ѧ����ʱ�������ۡ���������������������������ֹ����ѭ���ļ�����Ca2+Ũ�ȵ�ͻȻ���ߺͿ����½������������ġ�
�������������������˷�-��������
��������������£�������������֧���������崫�����˷ܳ嶯��Ӱ���½��������ģ�ֱ�����˹��̼���������֧��Ĺ�������Ҳ�����������������ۺ���������̼�����������֮ǰ���������ڼ�ϸ��Ĥ������һ���ɴ����Ķ�����λ��Ȼ��ų��ּ�ϸ����������Ӧ������������Ĥ�ĵ�仯Ϊ�������˷ܹ��̺��Լ�˿�Ļ���Ϊ��������������֮�䣬��Ȼ������ij���н��Թ��̰�������ϵ��������һ���̣���Ϊ�˷�-����������Ŀǰ��Ϊ�������ٰ���������Ҫ���裺���˷�ͨ�����ϵͳ����ϸ������������ܽṹ������Ϣ���ݣ������������ݹ�ϵͳ����Ca2+�ͷź��پۻ���
�������ϵͳ��������ϸ�����˷�-����������ʮ�ֱ�Ҫ�ġ��ú��и��͵ĸ�������Һ���ݼ���һ��ʱ�䣬�ٰ����Żص�һ������Һ�У������Ĵ�������ѡ���Ե��ƻ���ϸ���ĺ��ϵͳ����ʱ����ٸ���������Ӵ̼�����Ȼ�Կ�����õļ�ϸ��Ĥ����������λ��������������ϸ��������������֤�������Ĥ��һ�㼡ϸ��Ĥ�����Ƶ����ԣ����Ǻ��ߵ��������֣������Ҳ���Բ�����Na+����Ϊ������Ĥ��ȥ������������λ����һ��ϸ��Ĥ���˷ܶ�����������λʱ����һ��仯�����Ű���ϸ���ڲ��ĺ��Ĥ���������뵽�����ܽṹ��ÿ����С�ڵĽ��ԡ�
����ʵ�ʲⶨ��֤�������ⰲ��ʱ�����е�Ca2+Ũ�ȵ���10-7mol/L������Ĥ��ʼȥ�����ĺܶ�ʱ���ڣ�������1��5ms�����ߵ�10-5mol/L��ˮƽ���༴����100��֮�ࡣ�������Ca2+�ɺζ������÷�����45Ca����Ӱ�ȼ���֤�������ⰲ��ʱCa2+��Ҫͣ���;ۻ���z�߸������൱�ڼ���������ĩ�ز�λ����������ʱ��Ca2+������������ɢ����������ѭ��������������Ϊ������Ĥ�ϵĵ�仯�غ��ϵͳ���������ܲ���ʱ��һ����ij�����ӰѺ��Ĥ�Ϸ����ı仯���ݸ�����Զ�ļ�����Ĥ�ϵ�����Ca2+ͨ���Ľṹ��������߷��ӵı乹���ã�ʹͨ�����ţ����Ǽ������ڸ�Ũ�ȵ�Ca2+�Ͳ�����ܶ�������ɢ���뼡�������A˿����������һ�źŵ����ӣ�������Ϊ�Ǻ��Ĥ�ϴ��ڵ�һ�����⵰�ף�ƽʱ�Լ�����Ca2+ͨ�������л�е�������ã����ں��Ĥ�е�仯ʱ�����乹���ã�ʹԭ���Ķ������ý����Ҳ������Ϊ���Ĥ�����仯�������˵ڶ���ʹ������IP3�������µڶ��ڣ����ɺ���������Ca2+ͨ��ʹ֮���š��������������йص�Ĥ��Ĥ�е����ʼ���������Ӵ��������һ�ֿ�����ʽ�����п��ܵġ�
�����ͷŵ������е�Ca2+������Ѹ�ٳ�ȥ��Ŀǰ��֤�������ڼ�����Ĥ�ṹ�д��ڵ�һ�����������ת�˵����ʼ��Ʊû�Ľ�����Ʊ���һ��Ca2+����ʽATPø��Ŀǰ�ѱ������ᴿ����ռ������Ĥ������������60%���ڼ�����Ca2+��������ϣ������Էֽ�ATP�����������Ca2+����Ũ�Ȳ��������ɼ���ת�˵���������ǻ��ȥ�����ڼ�����Ca2+Ũ�ȵĽ��ͣ��ͼ��Ƶ���ϵ�Ca2+Ҳ���룬���������š�
�����ġ��������������ⲿ���ֺ���ѧ����
���������������ڵĹ��ܣ������������ܴ̼�ʱ�ܲ������̻ͣ��������������������˶��ͣ��ֿ����������á�������˷�ijһ���������̣����������̶�ǣ��ijһ����ʱ������������һ�����Ļ�е��������ֵ���������˷����������ɣ��ͼ������̳��ȵij˻������������ٶȳ��Ը��ɣ���ó������������ʡ�������������ʱ�����Բ�������Ϊ��������Ϊ�����Լ�����ʱ�������ٹ�����Ҫ����������ʱ�������ĸ��������ͼ��Ȿ���Ĺ���״̬��
�������������ڻ�ʵ�������¿��������ĸ�����Ҫ�����֣�һ�����ڼ�������ǰ�ͼ��ڼ����ϵģ����һ������˳�����ļ�ԭ��ά�����з����������������϶˹̶���������һ������һ��������������߾���ǰ���ɡ�ǰ����ʹ����������ǰ�ʹ���ij�̶ֳȵı�����״̬��ʹ������һ���ij��ȣ����Ϊ�����ȣ���������ǰ���ɵIJ�ͬ��ͬһ�����Ҫ�ڲ�ͬ�ij����������½�����������һ�ָ��ɳ�Ϊ�ɣ������ڼ��ʼ����ʱ���������ĸ��ɻ��������������Ӽ���ij����ȣ������谭����ʱ��������̡��������⣬����ijһ����ļ�����˵��ʵ�������Ӹ��ɡ��ر���ǰ���ɲ�Ӧ��������Ϊ�����ڼ�������ǰ�Ϳ�������ȵ�ǣ�������˼��Ȿ���Ľṹ�����ںɣ����ڴ�һ���̶Ⱦ����Եֿ��������������������������������ⲻ�ٱ������̣����ֵȳ��������༴��ʱ�������������������δ�г��ȸı䣻����������¼������Ӻɣ���Ȼ����Լ����������ʲôӰ�졣
��������������Ӱ�켡������ʱ��������������ѧ���ֵ�������������������ǰ���ɡ��ɺͼ��Ȿ���Ĺ���״̬��������������������Ҫ����ijһ����Ӱ�����취������ʹ�������ر�����ijһ�㶨ֵ���ı�Ҫ�۲����ص�ֵ���õ�һ�����ݲ�����һ���������������з�����
������һ��ǰ���ɻ�������ȶԼ���������Ӱ�엗����-��������
����Ϊ�˱�����ʵ������м��Ȿ���Ĺ���״̬�������ֲ��䣬ͨ��ѡ�ô�л�ٶȽ����������������賦��콳����ʵ�飬ʵ�鲼����ͼ2-24A��ʾ���������·����̶������ұ���һ����������������������¼��������ǰ����������������������������Ϸ���һ�����ƶ��İ�ť�����������ƶ����ı伡��ij����ȣ������۳����ȹ̶���ʲô���ȣ�ͬ��ť�����Ĺ̶����Dz��ܶ��ģ������ζ�ŰѺɹ̶�������ʱ��λ�ã�����������ʱ���������̶�ֻ�ܲ�����������ǰ����˵�ĵȳ������������ǾͿ��Թ۲�����Ȳ�ͬʱ��ͬһ�������ܲ���������Ӱ���ˡ�
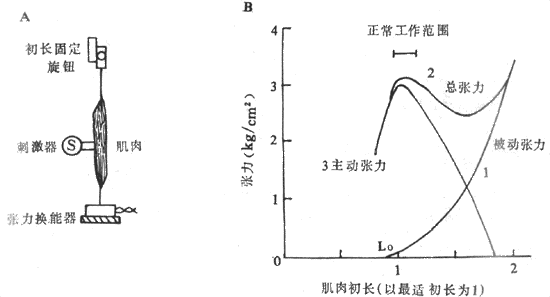
ͼ2-24 ��������ȶԼ���������Ӱ��
A����ʵ�鲼���У��·�����������������λ�ù̶����ܰ���������ת��Ϊ��Ӧ��
���źţ��Ϸ���ť�ɽ���������ڸ�����̼�ǰ�̶���Ԥ������
B��3�����߷ֱ���������ڳ����ȸı�ʱ�������ı��������������ָ�ı��������δ������
����������ı䣬�������������б��������Ļ����ϼ�������ʱ����������������ǰ��֮��
����ͼ2-24B�ij���-�������߷�ӳ�������θı伡��ij�����ʱ�������꣩�������������ϼ�¼���ļ������������������������꣩������1��ֻ�ı伡������Ȳ����̼���������ʱ�������ܵ���������Ϊ�����������ߣ�����ӳ�����������ij�ֵ��ԣ����ܵ�ǣ��ʱ����ij�ֻص�������ǣ������ij�̶ֳȣ��ﵽ�˵����ȣ������������������п��������֯���������������һ������ʱ���ơ�����2�Ǽ����ھ��в�ͬǰ���ɼ��Ѿ��б��������������½���һ������ʱ��¼���������仯�����ߵ�ÿһ�㶼�����Ǹ�������ʱ�������еı�������������ʱ�²���������֮�ͣ����������߳�Ϊ���������ߣ���ˣ�������2�����IJ�ͬ������ʱ����������ȥͬһ������ʱ�ı������������ܵõ�����3������ʾ�����ڲ�ͬǰ����ʱ�����������ܲ������������ʳ�Ϊ�����������ߣ�����ӳ�˱�ʵ����Ҫ�۲�����ݣ�����ͬǰ���ɻ�����ȶԼ����������ܲ���������Ӱ�죺��ǰ���ɿ�ʼ����ʱ��ÿ����������������������Ҳ��Ӧ�������ڳ���ijһ�Ⱥ�������ǰ���ɷ���ʹ��������Խ��ԽС��������Ϊ�㣬������3�Ҷ���ʾ���������Ҳ���Ա���Ϊ�����ڼ����ڵȳ�������������������������������С��������һ������ǰ���ɺ���֮���Ӧ�����ʳ����ȣ��൱��ͼ2-24B��������Lo��λ�ã��������ij���������½����������������������
�������������ʳ����������½������������ܲ��������������������ݼ��ⱻǰ��������ʱ��ÿһ��С���д֡�ϸ��˿���ϵ�ĸı������͡���֪������������������̣������Ǵּ�˿����ĺ��ź�ϸ��˿֮�������ã���������ȵĴ�С��������ÿ����С�ڵij��ȣ��༴ϸ��˿�ʹּ�˿�ص��ij̶ȣ��������־����ڼ�������ʱ�ж��ٺ��ſ����븽����ϸ��˿����á��������Ϸ������ּ�˿�ij�����1.5��m������M�������Ϊ0.1��m�ķ�Χ������ʱû�к��ţ������M�������к��ŵĴּ�˿���ȸ�Ϊ0.65��m��������ÿ��ϸ��˿���밵��0.65��m����δ��������ϸ��˿����Ϊ0.35��m�����༴��С���ܳ���Ϊ2.2��mʱ���ּ�˿�ϵ�ÿ�����Ŷ�����ϸ��˿���ã��������ʱ�ܳ�����ѵ�Ч���������������ǰ���ɻ����ʳ�����ʱ��ÿ����С�ڵij�������2.2��m����ͼ2-25��ͷ3��ʾ��������Լ���ǰ����ʹ��С�ڳ���2.0��m����ͷ2��������ÿ��ϸ��˿�ֶ����밵��0.1��m����ʱ����ϸ��˿����������������һ�����Ǵּ�˿����������IJ��֣������������ʱ�����õĺ�����Ŀ��δ���ࣨ�൱��ͼ2-25�еļ�ͷ2���������ټ�С��С�ڵij��ȣ���ϸ��˿���ܴ���M�����༡˿��غϺ;��������������������½���ͼ�м�ͷ1������֮�����ǰ���ɳ�������ǰ���ɣ�����ǰ��С�ڵij��Ƚ�����2.2��m��ϸ��˿�ʹּ�˿��غϵij̶���С��ʹ�ü�������ʱ�����õĺ�����Ҳ���٣�����������������½�����ǰ����ʹ��С�ڳ������ӵ�3.5��mʱ��ϸ��˿��ȫ���ɰ�����������ʱ�����ܴ̼�ʱ���ٲ�������������ͼ�м�ͷ4�����ɴ˿ɼ���ͨ��ǰ���ɶԼ�С���д֡�ϸ��˿�غϳ̶ȵ�Ӱ�죬����˵������������-�������ߵ��ص㡣

ͼ2-25 ��ͬ������ʱ�֡�ϸ��˿�غϳ̶ȺͲ��������Ĺ�ϵʾ��ͼ
�����ü�С���ڲ�ͬǰ����ʱ�֡�ϸ��˿���λ�õĸı䣬��˵����ͬǰ����ʱ�����������������IJ�ͬ���ڼ�ͷ1��ָ�ij�����ʱ��ÿ����С��������ϸ��˿���밵�����࣬�����ص���������������������ż������ã��ڼ�ͷ2��3��ָ������£���С����ȫ�����Ŷ�����ϸ��˿����ã���������������������ڼ�ͷ4������£�ϸ��˿�ȫ���ɰ�����������ʧȥ��������������
ͼ2-26 ���������-�ٶȹ�ϵ����
�ڼ���ǰ���ɹ̶����ʵ�ֵ�������£��ı�ɶԼ�����������������꣩
���ٶ��ٶȣ���������꣩���ϵ��Ӱ�죬���Ϊ����-�ٶȹ�ϵ���ߣ�����1����
�ɴ����߿��������ͬ��ʱ��������ʣ��Ҳ������꣩�����������2
��������������ɶԼ���������Ӱ�죭����-�ٶ�����
������ǰ��������ʵ����װ�õ������ʹһ����������ǰ���ɹ̶������������Ϊ�ظı�ɣ����ɹ۲첻ͬ�ɶԼ���������Ӱ�졣һ������£����Ѽ����ǰ���ɹ̶�����������ǰ���ɣ���ʱ���ֵı���������С����ͼ2-24B����Ȼ������θı�ɵ�����¹۲켡������ʱ��������������κ�ǰ���ɵ�����£�������Ӻɳ����˼�������ʱ���ܲ��������������ע�⸺�ɵ�����ֵ�ͼ��������������ֵ��������ͬ��������λ��������������ͬ��ֵ������ô��������ʱ��ֻ���������������ּ��ⳤ�ȵĸı䡣��ˣ��ڸı�ɵ�ʵ�������ӵĺɶ�ӦС����������������ô����������ʱ����������������������ɵ�ֵʱ�����������һ���̶ȵij������̣�ʹ�ƶ�����ͬ�ľ��룬�������ں���Ҳ�������һ�������ٶ����������dz��ٶȻ�ƽ���ٶȣ�������С���������������������س���������ɣ����ҳ��ֽϴ�����̳��Ⱥ������ٶȣ����൱�ڸ���ֵ�ļ�������ȴ�����̵Ĺ����б��ֲ��䡣�����͵õ��˸ı��ʱ����������������������ٶȱ仯�Ĺ�ϵ���ߣ�Ϊ�˼���������ʣ�һ��ֻ���������ٶȺ������Ĺ�ϵ������ͼ2-26����1����Ϊ����-�ٶ����ߡ�����������һ��˫���ߣ��������ʾ�������������������������ʾ�����ٶȣ�˫���ߵ�������˵������ߴ��³ʷ��ȵĹ�ϵ�����ɼ�Сʱ��ʹ���������������С�����ɵõ�һ���ϴ�������ٶȣ�������ͬ�����ཻ�ĵ㣬˵����������Ϊ��ʱ�����Եõ��ü����ڵ�ʱ�Ĺ���״̬�µ���������ٶȣ���ͼ2-26����V0��ʾ������ʱ�����ɣ����Ⲣδ�����������������������ͬ�����ཻ�ĵ㣬�ɵ�ֵ�൱�ڼ������ܲ����������������ʱ�����ƶ����ɣ�Ҳû���������������������������֮�䣬�ڲ�ͬ�ĺ�ʱ���ܿ��������ڲ����븺����ͬ�������������ʹ�����ƶ�һ�����룬�������͵���������Ϊ���������������������й�����������Ժ��൱�����������30%����ʱ�������������������ͼ2-26������2��ʾ��
�����������������������ĸı�Լ���������Ӱ��
����������ǰ���ɵĸı�Լ�������ʱ���������������ٶ��Լ�������������ѧ���ֵ�Ӱ�죬��Ȼ���ڼ����״̬�㶨������¶��������������ı������IJ�ͬ��Ӧ���������״̬Ҳ�ǿ��Ըı�ģ���Ҳ����Ӱ�켡��������Ч�ʡ����磬ȱ�������ж�����������Դ����ȱ�����Լ�����ԭ��������˷�-�������������⵰���ʻ���Ź������Եĸı䣬�����ܽ��ͼ���������Ч�����������ӡ��������������ص���Һ���������ͨ��Ӱ�켡����������ƶ�����������Ч������Ӱ�켡������Ч���ļ����ڲ�����״̬�ĸı䣬����Ϊ��������������contractility���ĸı䣬�������ڼ�������ʱ�ⲿ������ǰ���ɸı������µ�����Ч���ĸı䡣������������Ȼ�ڸ����ϱȽ����ף����ھ��������Ҫ������Щ�ı������ڼ������������ĸı���������Щ�����ڸ��������ĸı������𣬳���ʮ�����ѡ����磬һ������������������ˣ����������ڼ���������������ߣ���Ҳ�������������������ǰ���������ʳ����ȣ�һ�������������ʱ�������ٶ������ˣ����������ںɵļ�С��Ҳ���������������ʳ�������Ҳ���������ڼ���������������ߣ����������֮�������˵�����Ѽظ��ݼ���ij����ѧָ��ĸı䣬ȷ���Ƿ����˼������������ĸı䡣�������Ͻ����������������ĸı�Լ��������ĸ���ѧ���ֵ�Ӱ���ǡ���ѡ���ԡ��ģ���������������߿�ʹͼ2-24B�еij���-�������ߣ�������������ͼ2-26�е�����-�ٶ����ߵ�λ�ö����ߡ���Ȼ��Ϊ�˼�����������Ƿ�ı���ٻ���һ��������������ʮ�ָ��ӵģ�Ϊ�˼�㣬�������ͬһ����������ǰ������������������м��������ٶȵĸı䣬��ʹ����ά�����ʳ����ȶ�����������ĸı䣬��ʾ�����������������˸ı䣻��Ϊ����Щ�����¿��Կ϶�����Щ����Ч���ĸı䲢��������ǰ���ɵĸı����������ֻ�������ڼ�����������ܵĸı�����ġ�
�������ģ�����ĵ������͵������ĸ���
�����������������ϸ���ܵ�һ�ζ̴ٵĴ̼�ʱ�����Dz���һ�ζ�����λ�������ų���һ�λ�е���������߳�Ϊ����������������ʱ���������ĸ���������ͬ�������������ǵȳ��ģ�Ҳ�����ǵ��ŵġ�ǰ�������ļ�������ʱ������ѧ���֣������Ե�����Ϊ�۲��������з����ġ������������ڣ������������˶���֧���½�����Ȼ����ʱ��������������ؽ��������������̼�������б�Ҫ��һ�������������ܵ���ͬƵ�ʵ������̼�ʱ���ܷ����������Ϊ�˱��ڷ������ȹ۲�һ�¼��ⵥ����ʱ��仯�ͻ�е�仯��ʱ���ϵĹ�ϵ��ͼ2-27��è��ǰ��һ�εȳ�������ʱ�����仯��ȫ���̣�ͬʱ��¼�˼���Ķ�����λ��ע��ͼ�е練Ӧ�Ŀ�ʼҪ���������ӵĿ�ʼΪ�磬���ҵ�仯�������ﵽ������ǰ���ѽ�������������ߵ�Ϊ�磬����ȫ���̿ɷ�Ϊ�����ں������ڣ�ǰ�߳���ʱ��Ϻ���Ϊ�̡�������������ʱ�����ⲻͬ�����������죬���˵����⼡��һ�ε�����������10ms�����賦���ɴ�100ms���ϡ�

ͼ2-27 è��ǰ���ĵȳ�����������
M����������ʱ�������仯����
E�������˫�ද����λ��¼ʱ��ÿ ���൱��0.01s
������������������������̼�������������������̼���Ƶ�ʶ��в�ͬ����ͼ2-28��ʾ���ڴ̼���Ƶ�ʽϵ�ʱ����ÿһ���µĴ̼�����ʱ��ǰһ�δ̼�����ĵ��������̣����������ڣ��Ѿ�����������ÿ�δ̼�������һ�ζ����ĵ����������̼�Ƶ�����ӵ�ijһ��ʱ�������Ĵ̼��п�����ǰһ�������������ڽ���ǰ�����A�⣬���Ǽ����������д���һ���̶ȵ����̻��������ڵĻ����Ͻ����µ���������������ν�������̵ĸ��ϣ���������������ȥ������ͱ���Ϊ����ȫǿֱ���������ص���ÿ���µ�������������ǰ�������������ڹ����У�������������γɾ���Σ�����̼�Ƶ�ʼ������ӣ���ô������п�����ǰһ�������������ڽ�����ǰ���������ڵĶ��㿪ʼ�µ����������Ǹ��������������ȱ仯�����ں϶�����������ʹ��������ϵľ������ʧ���������ȫǿֱ������

ͼ2-28 ��ͬƵ�ʵ������̼��Թ�����������Ӱ��
ÿ�������У��Ϸ������Ǽ���ȳ������ļ�¼���·������Ǽ������λ�ļ�¼��
ע�����϶��µ��̼�Ƶ����������ʱ����е�����ɳ������ںϣ�
���ֲ���ȫ����ȫǿֱ��������������λʼ�ձ˴˷��룬�������ںϺ͵���
�������������������˶��������������˷ܳ嶯���ǿ��������ģ����ڹ���������������������ȫǿֱ������ֻ����ǿֱ�����ij���ʱ��ɳ��ɶ̡�ǿֱ������Ȼ���Բ������������Ч�������磬ǿֱ�������ܲ�������������ɴﵥ������4�����ҡ�������Ϊ������ֻ����һ�δ̼�ʱ���ͷŵ������е�Ca2+�ܿ챻�������ϵ�Ca2+�û����뼡�������������̼���ʹ�����е�Ca2+ά����һ�����͵ĸ�Ũ��ˮƽ����ͬ���ⵥ�����ij���ʱ�䲻ͬ������������������ȫǿֱ����������ٽ�Ƶ���ڲ�ͬ����Ҳ��ͬ�����磬���������ٵ�������ֱ����Ҫÿ��Լ350�εĸ�Ƶ�̼����ܲ�����ȫǿֱ�����������������ı�Ŀ�㼡ֻ��ÿ��Լ30�ε�Ƶ�ʾ��ˡ��������ڲ���ȫǿֱ��������ȫǿֱ����������ÿ�δ̼����ֵļ������λֻ����Ƶ�ʼӿ죬ȴʼ�ո��Է�������ᷢ���ںϻ��ܺͣ��������ڼ���Ķ�����λֻ����1��2ms�����̼�Ƶ�ʼ��ٵ���һ�δ̼�����ǰһ�δ̼�������Ķ�����λ�����ڼ�ʱ����֯�����ô����˷ܵľ��Բ�Ӧ�ڣ���ʱ�µĴ̼�����Ч���Ȳ��������µĶ�����λ������Ҳ�������µ�������
�����塢ƽ�����Ľṹ����������
����ƽ�����㷺�ֲ����������������������Լ�Ѫ�ܺ�������ֳ��ϵͳ������������ͬ������ÿ������ά������ϸ���������˶�ͨ������ͬ����������ƽ����ϸ���������ӣ��γɹ�״�ṹ���п����٣��ڹ����Ͽ���ͨ�����̺Ͳ�������ʹ���ٷ����˶��ͱ��Σ�Ҳ�ɲ������������������������ʹ���ٶԿ����Ӹ��ɶ�����ԭ�е���״��ǰ����θ�ͳ��������綯��Ѫ�ܡ���Լ���ȡ����⣬Ҳ�������ڹ��������ļ��������ѷֲ��ڲ�ͬ���ٵ�ƽ��������������ͬ�������Ժ͵��ڻ��Ƶ���֯��������Щ���ٵ�ƽ�����к�����һ�����Զ������ԣ���Щ���������������ֻ����֧����������ά���嶯����ʱ�ų�����������������������֮�䣬�������Ÿ��ֵĹ�����ʽ����ʹƽ�����ķ������ѡ�
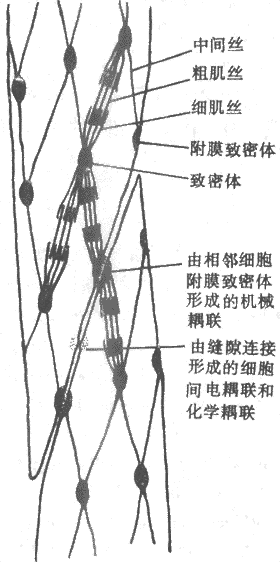
������һ��ƽ������ϸ�ṹ����������
����ƽ������ȻҲ����ͬ���������Ƶļ�˿�ṹ�����������Dz����������������ƽ�ж���������У�ƽ�����ļ�˿�����Լ��ġ�����ġ����У���ͼ2-29�Ǹ�������۲������ƽ����ϸ���ڲ�����ṹͼ�������ص���ϸ���ڲ�����һ��ϸ���Ǽܣ�����һЩ��Բ�εij�Ϊ������Ľṹ������Ҳ����س�����ϸ��Ĥ���ڲ࣬��Ϊ�����������Һ���������ϸ�������ƽṹ��ԣ���������ϸ��ĤҲ�ڴ˴����������������ͬ�����һ�ֻ�е���������������ϸ���������Ĵ��ݣ�ϸ����Ҳ���ڱ��������ʽ�����϶���ӣ����ǿ���ʵ��ϸ����ĵ������ͻ�ѧ����������������������з�����ͬ������Z�������Ƶĵ��׳ɷ֣�����Ϊ�����ֽṹ��������ϸ��˿���ӵIJ�λ�����⣬���������������֮�仹��һ��ֱ�����ڴ֡�ϸ��˿֮���˿״����ڣ�������һ�ֳ�Ϊ�ᵰ�ף�desmin���ľۺ��塣������˿״�������������������Ĥ�ڲ�����������γ���������ϸ���ڹ��ܡ�
����ƽ����ϸ���е�ϸ��˿��ͬ���������Ƶķ��ӽṹ�����������Ƶ��ף�ͬһ�����ƽ�����������˵������ǹ�������2�����Ʋ�ƽ�����������д���ϸ��˿���ڣ����ǵ����д�����ϸ������ƽ�С�����෴�������м���������ȴֻ�й�������1/4�������������������ϵ�3��5��ϸ��˿�ᱻ������Ŀ�ּ�˿���ƣ��γ������ʽ�����У�����ܾ��������ڹ������м�С�ڵĹ��ܵ�λ��
ͼ2-29ƽ�Ǽ��ڲ��ṹ�����ϵģʽͼ
����һ��ƽ����ϸ�������Σ�ֱ��2��5��m���䳤�ȿɱ��Ժܴ�Լ����Ϊ400��mʱ�Dz������������ʳ��ȡ�����û�й����������ļ�����������ļ���ϵͳ����ϸ��Ĥֻ��һЩ�������еĴ�״���룬���书���в������������ʹ��ϸ��Ĥ�������ϸ�����֮�ȸ�Ϊ�Ӵ���˺ͼ�˿�����IJ��Ǻ�ܻ���ϵͳ�����Ǽ�Ĥ��ϸ��������ʱ��ϸ����Ca2+����Ĥ�ڣ���ƽ����ϸ���п���Ĥ�ļ�����Ҳ������ϸ����Ca2+����⡣һЩ�˷��Ե��ʡ����ػ�ҩ��ͬ��Ĥ������ʱ��ͨ��G-�����ڰ����в����ڶ���ʹ������Ca2+���е�Ca2+�ͳ�����ƽ������ϸ��˿�в����ڼ��Ƶ��ף����Ca2+����ƽ����ϸ���д֡�ϸ��˿����еĺ���ѭ���Ļ������������ͬ��Ŀǰ��Ϊ�����ŵļ��ʼ���������ữ������������һ��Ϊ��������ø�Ļ���������Ca2+�Ƚ���ڰ�����һ�ֳ�Ϊ�Ƶ����ף�calmodulin�������⵰���ʣ����߽��4��Ca2+֮���ʹ��������ø���ʹATP�ֽ⣬�ɴ˲��������������ں��Ų�ʹ���Ŵ��ڸ�����״̬������ƽ��������ƽ�������ż���Ļ�����Ҫ�ϳ���ʱ�䣬���ƽ���������Ļ�����һ�¡�
����������ƽ�����ڹ����ϵķ���
�����������ڸ���������ƽ�����ڹ����������б�ܴ�һ��ɷ�Ϊ�����ࣺһ���Ϊ�λ��multi-unit��ƽ����������������ƽ����ϸ���ڻʱ���Զ��������ƹ�����ϸ��������ë������Ĥ����˲Ĥ����è�����Լ���Ѫ��ƽ�����ȣ����Ǹ�ϸ���Ļ��������֧�������ɢ����ϸ���ļ��ص�Ӱ�죻��һ���Ϊ��λ��single-unit��ƽ�����������ļ���֯�����и�ϸ��ͨ��ϸ����ĵ����������Խ���ͬ���Ի������ƽ�������������ԣ���û��������֧��ʱҲ�ɽ��н����������������������ϸ���������Ժ������Ե����ã�����θ�����ӹ��������ƽ����Ϊ����������һЩƽ����������������ص㣬���ѹ�����һ�࣬��С������С����ƽ����һ����Ϊ���ڶλƽ�����������������ԣ�����ƽ����û�������ԣ���������ǣ��ʱ����Ϊһ��������Ӧ����Ҳ���뵥λƽ������
����������ƽ������Ŀ��ƺ͵���
������ƽ�������������Ծ��ж�����һ�������ǻ���ܵĵ���Ҳ�Ƕ��ֶ����ģ�������������������������ƽ����������֧�䣬��������������ϵͳ��������֧�䣬���г�С����һ��ֻ���ܽ���ϵͳһ��������֧���⣬�������ٵ�ƽ����ͨ�����ܽ��к�����������֧�䡣ƽ������֯���ر���������ƽ���������л��������Դ��ڣ����߽���������Ӱ�죬�����л������оֲ���������Ԫ������������ַ��䡣ƽ��������-����ͷ��Щ���ƹ��������������к�����������ṹ��ʽ��֧��ƽ��������������ά�ڽ������֯ʱ��η�֧����֧��ÿ��һ���������һ���������״����Ϊ�����壬���к��з������ݣ��������嶯����ʱ�����ͷ����е��ʻ������������ʣ�ÿ��������Ͱ�ϸ���ľ�����̶���ƽ��ԼΪ80��100nm����˵������ĩ���ͷų����ĵ��ʷ���Ҫ��ɢ��Զ������ܴﵽ��ϸ��������ϸ������ĩ�ҵĹ�ϵҲ�������ǹ̶��ģ����ǵ��ʷ��ӿ��Ե�����־��иõ��������ƽ����ϸ���������ܽ���������Ӱ�졣
����ƽ����ϸ��Լ���֣�55����60mV�ľ�Ϣ��λ���������ƺ��������ơ���λƽ����ϸ���в���������λ������������ͨ��ϸ����ͨ����ʹ����ϸ��Ҳ����������λ��
���������